



Schistosomiasis is a parasitic disease of cattle and produces visceral or nasal disease in dairy cattle and work bullocks. Clinical signs include diarrhea, weight loss, and anemia. Lesions include granulomas in the intestines and liver. It is easily diagnosable by signs and is treated with anthiomaline or praziquantel.
Schistosomiasis is common in cattle and is rarely seen in other domestic animals in Africa and Asia. Although Schistosomes may act as important pathogens under rare conditions favoring intensive transmission, most infections in endemic areas are subclinical. However, high prevalence rates of subclinical infections cause significant losses due to long-term effects on growth and productivity and increased susceptibility to other parasitic or bacterial diseases. Market values of cattle, especially those affected with snoring disease, can be severely reduced.
Schistosomes are members of the genus Schistosoma, family Schistosomatidae. Adult worms are obligate parasites of the vascular system of vertebrates. Schistosomes are dioecious. The mature female is more slender than the male and normally is carried in a ventral groove, the gynecophoric canal, that is formed by ventrally flexed lateral outgrowths of the male body.
Parasitic Schistosomes of Ruminants
Type of Schistosomes | Geographic Distributions |
|---|---|
S mattheei | Southeastern Africa, from the Cape Province in South Africa northward to Tanzania and Zambia |
S bovis | Mediterranean region and Middle East, and is common in northern, western, and eastern Africa (except Egypt) extending southward to Central Angola, southern Congo, and possibly northern Zambia |
S curassoni | Senegal, Mauritania, Mali, Niger, and Nigeria |
S spindale | India (Karnataka, Maharastra, Andhra Pradesh, Madhya Pradesh, Tamilnadu and Assam), Sri Lanka, Indonesia, Malaysia, Thailand, and Vietnam |
S indicum | Indian subcontinent, Bangladesh |
S nasale | India, Sri Lanka, Bangladesh, and Myanmar |
S incognitum | India, Thailand, and Indonesia |
S japonicum | Endemic in several countries of the Far East |
Of the 19 species reported to naturally infect animals, 8—all parasites of ruminants—have received particular attention, mainly because of their recognized veterinary significance: Some of these species are known to interact in areas where they coexist, and instances of interspecific hybridization have been reported, eg, the cattle parasites S bovis and S curassoni. Novel molecular tools have also provided evidence for the natural hybridization between S haematobium, a parasite of people, and S mattheei, S bovis, and S curassoni. The hybridization between human and ruminant schistosomes is of particular interest, because for this to occur, host switching must have taken place of S mattheei, S bovis, or S curassoni into people or of S haematobium into domestic livestock.
To differentiate the various Schistosoma spp, egg morphology (size, shape) is used. The species can also be differentiated through such taxonomic features as morphologic (adult worms), life-cycle, or behavioral characteristics; chromosomes; host specificity; or enzyme and DNA studies.
Life Cycle, Transmission, and Epidemiology of Schistosomiasis in Cattle
Schistosomes live in the mesenteric and hepatic veins of the host (except for S nasale, which lives in the nasal veins), where they feed on blood and produce eggs with a characteristic terminal or lateral spine. Eggs passed in the feces must be deposited in water if they are to hatch and release miracidia, which invade suitable water snails (Bulinus, Biomphalaria, Indoplanorbis, and Lymnaea spp) and develop through primary and secondary sporocysts to become cercariae. When fully mature, the cercariae leave the snail and swim freely in the water, where they remain viable for several hours. Ruminants are usually infected with cercariae by penetration of the skin, although infection may be acquired orally while animals are drinking. During penetration, cercariae develop into schistosomula, which are transported via the lymph and blood to their predilection sites. The prepatent period varies according to the species but is generally 45–70 days.
The occurrence of cattle schistosomes within their range is discontinuous, depending on the presence of intermediate snail hosts, their level of infection, and the frequency of water contacts. In areas where conditions are favorable, prevalence rates of infections in cattle may be 40%–70% and commonly higher.
The increased host range of the hybrid parasites and changes in host distribution seen in Africa may have a direct impact on transmission of these schistosomes. Laboratory hybrids have been observed to acquire enhanced characteristics such as infectivity, fecundity, and growth rates.
There is strong evidence that acquired immunity to schistosome infection in cattle exists. This immunity mainly acts through suppression of worm fecundity. Examination of naturally infected animals has shown that partial protection against reinfection also occurs, and acquired resistance to schistosomes is of major importance in the regulation of infection intensity in the field.
Clinical Findings and Lesions of Schistosomiasis in Cattle
Visceral Schistosomiasis
In the great majority of cases, visceral schistosomiasis in endemic areas is subclinical and characterized by a high prevalence of low to moderate worm burdens in the cattle population. Although few or no overt clinical signs may be recognized in the short term, high prevalence rates of chronic schistosome infections cause significant losses on a herd basis. These losses are due to less easily recognizable effects on growth and productivity, as well as increased susceptibility to other parasitic and bacterial diseases.
Occasional outbreaks of clinical intestinal schistosomiasis due to S mattheei, S bovis, or S spindale have been reported. They are usually restricted to young livestock and adult animals undergoing relatively heavy primary infections under conditions of intensive transmission. The disease is characterized by diarrhea, weight loss, anemia, hypoalbuminemia, hyperglobulinemia, and severe eosinophilia that develop after the onset of egg excretion. Severely affected animals deteriorate rapidly and usually die within a few months of infection, while those less heavily infected develop chronic disease with growth retardation.
In the intestinal and hepatic forms, adult flukes are found in the portal, mesenteric, and intestinal submucosal and subserosal veins. However, the main pathologic effects are associated with the eggs. In the intestinal form, passage of eggs through the gut wall causes the lesions, while in the hepatic form, granulomas form around eggs trapped in the tissues. Other hepatic changes include medial hypertrophy and hyperplasia of the portal veins, development of lymphoid nodules and follicles throughout the organ, and periportal fibrosis in more chronic cases. Extensive granuloma formation also is seen in the intestine. In severe cases, numerous areas of petechiation and diffuse hemorrhage are seen in the mucosa, and large quantities of discolored blood may be found in the intestinal lumen. Frequently, the parasitized blood vessels are dilated and tortuous. Vascular lesions also may be found in the lungs, pancreas, and bladder of heavily infected animals.
The hybridization events reported between animal and human schistosomes may result in phenotypic characteristics that influence pathology (and drug sensitivity).
Nasal Schistosomiasis
Nasal schistosomiasis is associated with cauliflower-like growths on the nasal mucosa, causing partial obstruction of the nasal cavity and snoring sounds when breathing. Hemorrhagic and/or mucopurulent nasal discharge is a common feature. Adult flukes are found in the blood vessels of the nasal mucosa, but again, the main pathogenic effects are associated with the eggs, which cause abscesses in the mucosa. The abscesses rupture and release eggs and pus into the nasal cavity, which eventually leads to extensive fibrosis. In addition, large granulomatous growths are common on the nasal mucosa and occlude the nasal passages and cause dyspnea.

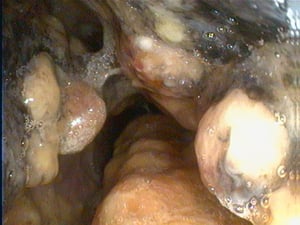
Cauliflower-like growth, which causes partial obstruction of the nasal cavity.
Cauliflower-like growth, which causes partial obstruction of the nasal cavity.
Courtesy of Dr. S. Yogeshpriya.
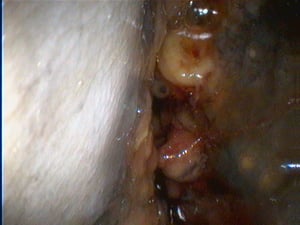
Obstructed nasal cavity in nasal schistosomiasis.
Obstructed nasal cavity in nasal schistosomiasis.
Courtesy of Dr. S. Yogeshpriya.

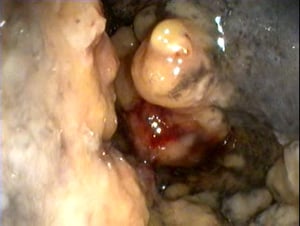
Nasal granuloma obstructing the nasal passage.
Nasal granuloma obstructing the nasal passage.
Courtesy of Dr. S. Yogeshpriya.

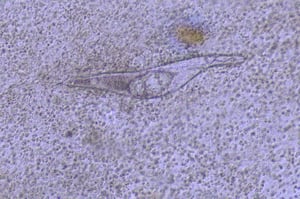
Fully developed miracidium in S nasale egg from nasal washing.
Fully developed miracidium in S nasale egg from nasal washing.
Courtesy of Dr. S. Yogeshpriya.

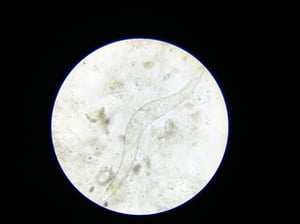
Fully developed miracidium inside the egg of schistosoma.
Fully developed miracidium inside the egg of schistosoma.
Courtesy of Dr. S. Yogeshpriya.

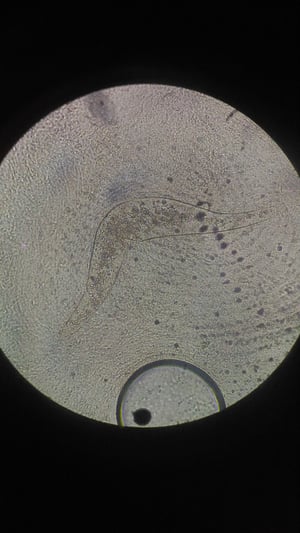
Boomerang-shaped S nasale egg from feces.
Boomerang-shaped S nasale egg from feces.
Courtesy of Dr. S. Yogeshpriya.
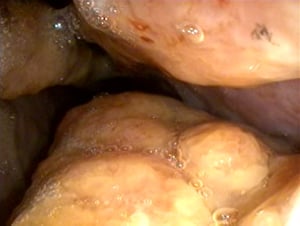
Nodular growths in the nasal cavity of a cow with schistosomiasis.
Nodular growths in the nasal cavity of a cow with schistosomiasis.
Courtesy of Dr. S. Yogeshpriya.

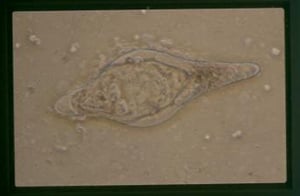
An embryonated egg of Schistosoma bovis as seen in a fecal sample from a cow. Note the characteristic spindle shape.
An embryonated egg of Schistosoma bovis as seen in a fecal sample from a cow. Note the characteristic spindle shape.
Courtesy of Francis McNeilly.
Cauliflower-like growth, which causes partial obstruction of the nasal cavity.
Cauliflower-like growth, which causes partial obstruction of the nasal cavity.
Courtesy of Dr. S. Yogeshpriya.
Obstructed nasal cavity in nasal schistosomiasis.
Obstructed nasal cavity in nasal schistosomiasis.
Courtesy of Dr. S. Yogeshpriya.
Nasal granuloma obstructing the nasal passage.
Nasal granuloma obstructing the nasal passage.
Courtesy of Dr. S. Yogeshpriya.
Fully developed miracidium in S nasale egg from nasal washing.
Fully developed miracidium in S nasale egg from nasal washing.
Courtesy of Dr. S. Yogeshpriya.
Fully developed miracidium inside the egg of schistosoma.
Fully developed miracidium inside the egg of schistosoma.
Courtesy of Dr. S. Yogeshpriya.
Boomerang-shaped S nasale egg from feces.
Boomerang-shaped S nasale egg from feces.
Courtesy of Dr. S. Yogeshpriya.
Nodular growths in the nasal cavity of a cow with schistosomiasis.
Nodular growths in the nasal cavity of a cow with schistosomiasis.
Courtesy of Dr. S. Yogeshpriya.
An embryonated egg of Schistosoma bovis as seen in a fecal sample from a cow. Note the characteristic spindle shape.
An embryonated egg of Schistosoma bovis as seen in a fecal sample from a cow. Note the characteristic spindle shape.
Courtesy of Francis McNeilly.
Diagnosis of Schistosomiasis in Cattle
Clinical signs, egg identification, and serology
Because signs and history alone are insufficient to distinguish visceral schistosomiasis from other debilitating diseases, diagnosis should be confirmed by the presence and identification of eggs in the feces of the infected animal. At necropsy, macroscopic examination of the mesenteric veins for the presence of adult worms or microscopic examination of scrapings of the intestinal mucosa or of crushed liver tissue (both for eggs) may prove easier.
Eggs of S bovis, S curassoni, and S mattheei are spindle-shaped. Because of the interspecific hybridization between S bovis and S curassoni and the natural hybridization between S haematobium and S mattheei, S bovis, and S curassoni, eggs of intermediate morphology may be seen. The eggs of S spindale are more elongated and flattened on one side, and those of S nasale are boomerang-shaped. The oval eggs of S japonicum are relatively small, with a rudimentary spine.
Very low fecal egg excretion is commonly seen in chronic infections; therefore, it may be preferred to use quantitative miracidial hatching techniques which, in addition to being more sensitive, also provide information on the viability of the eggs excreted in the feces.
Molecular analysis can help to identify an organism and its relationship to close phylogenetic relatives. During the past few years, several authors have tried to develop new diagnostic tools with higher specificity and sensitivity, some based on the detection of Schistosoma DNA by PCR.
Treatment and Control of Schistosomiasis in Cattle
Anthiomaline or praziquantel
Praziquantel (25 mg/kg, PO, in water buffalo; 30 mg/kg, PO, in cattle) is highly effective to treat schistosomiasis in cattle; however, two treatments 3–5 weeks apart may be required. However, for practical and economic reasons, schistosomiasis in domestic stock is rarely treated in certain areas. Only in China, where infected livestock constitute important reservoirs of human infection, have mass treatments with praziquantel been practiced widely. In India, all the affected cattle that are brought for veterinary attention are treated with anthiomaline or praziquantel. Triclabendazole showed adulticidal effect at a dosage of 20 mg/kg against female schistosome worms.
For nasal schistosomiasis, treatment is started with an injection of anthiomaline (lithium antimony thiomalate, 15 mL, IM). Affected cows respond after the first dose of anthiomaline, and there will be reduction in the size of the nasal granuloma. Anthiomaline injections can be repeated at weekly intervals, if needed, until further reduction in the size of nasal granuloma is achieved and snoring sounds are reduced. Two or three injections usually produce a complete recovery.
The most effective way to control cattle schistosomiasis in endemic areas is to prevent contact between the animals and the parasite by fencing of dangerous waters and supplying clean water. Unfortunately, this is not always possible in parts of the world where nomadic conditions of management prevail. Other methods of control include destruction of the snail intermediate host population at transmission sites, either by chemical or biologic methods, or their removal by mechanical barriers or snail traps. Ecologic measures against the snails that aim to render their habitat unsuitable for survival, such as drainage, removal of water weeds, and increased water flow, have also proved valuable. These measures not only help reduce the transmission of schistosomiasis but also help control other parasitic trematodes such as Fasciola gigantica and paramphistomes, which also have water snails as intermediate hosts and frequently are found in the same localities as schistosomes.
Key Points
Control of snails, avoiding animal grazing near snail-infested areas, periodic deworming in bullocks, and treatment at the early stage of infection will help in control of schistosomiasis in ruminants.
Transmission of infection in nasal schistosomiasis occurs by percutaneous penetration of cercaria of S nasale from infected Indoplanorbis sp snails.
For More Information
World Health Organization (WHO); Schistosomiasis. Geneva: 2014.





