



Tyzzer disease is caused by Clostridium piliforme. The disease affects a variety of animals, including mammals and birds. It is characterized by a triad of lesions that include colitis, hepatitis, and myocarditis, although the three are usually not present together in a single animal. Treatment is supportive and nonspecific, with a low success rate.
Tyzzer disease is caused by Clostridium piliforme and affects a wide range of animals in many regions of the world. It was first described in mice in 1917. Several years later, it was reported in laboratory rabbits and then in other small laboratory mammals, including guinea pigs, hamsters, gerbils, and rats. It is a highly fatal disease of young foals, but it is rare in other domestic animals, including dogs, cats, and calves. It has been reported, albeit rarely, in a variety of wildlife, including muskrats, cottontail rabbit, coyote, gray fox, lesser panda, snow leopard, raccoon, marsupials, white-tailed deer, and a few avian species.
The disease primarily affects young, well-nourished animals, especially those fed high-protein diets, during periods of stress. Some species appear resistant unless stressed or immunosuppressed, whereas others appear to be susceptible without immunosuppression. Dietary factors, including excessive nitrogenous diets fed to laboratory animals or to nursing mares, may cause immunosuppression and may predispose susceptible animals to the disease.
In all animal species, Tyzzer disease is characterized clinically primarily by liver failure and icterus, a consequence of hepatic necrosis. Less frequently, diarrhea is observed. A triad of lesions consisting of hepatitis, colitis, and myocarditis is considered classic for the disease. However, although hepatitis seems to be constant in all species, colitis and myocarditis occur sporadically in some species. For instance, colitis and/or myocarditis rarely occurs in horses.
Although little is known about predisposing factors, it is thought that immunosuppressive agents and drugs, and some antibacterials, especially sulfonamides, may predispose animals to the disease.
Diagnosis is usually confirmed by histopathologic analysis and supported by PCR assay of C piliforme. Treatment is mostly supportive with fluids, although foals may be treated with administration of penicillin; the rate of success, however, is low.
Etiology and Pathogenesis of Tyzzer Disease in Animals

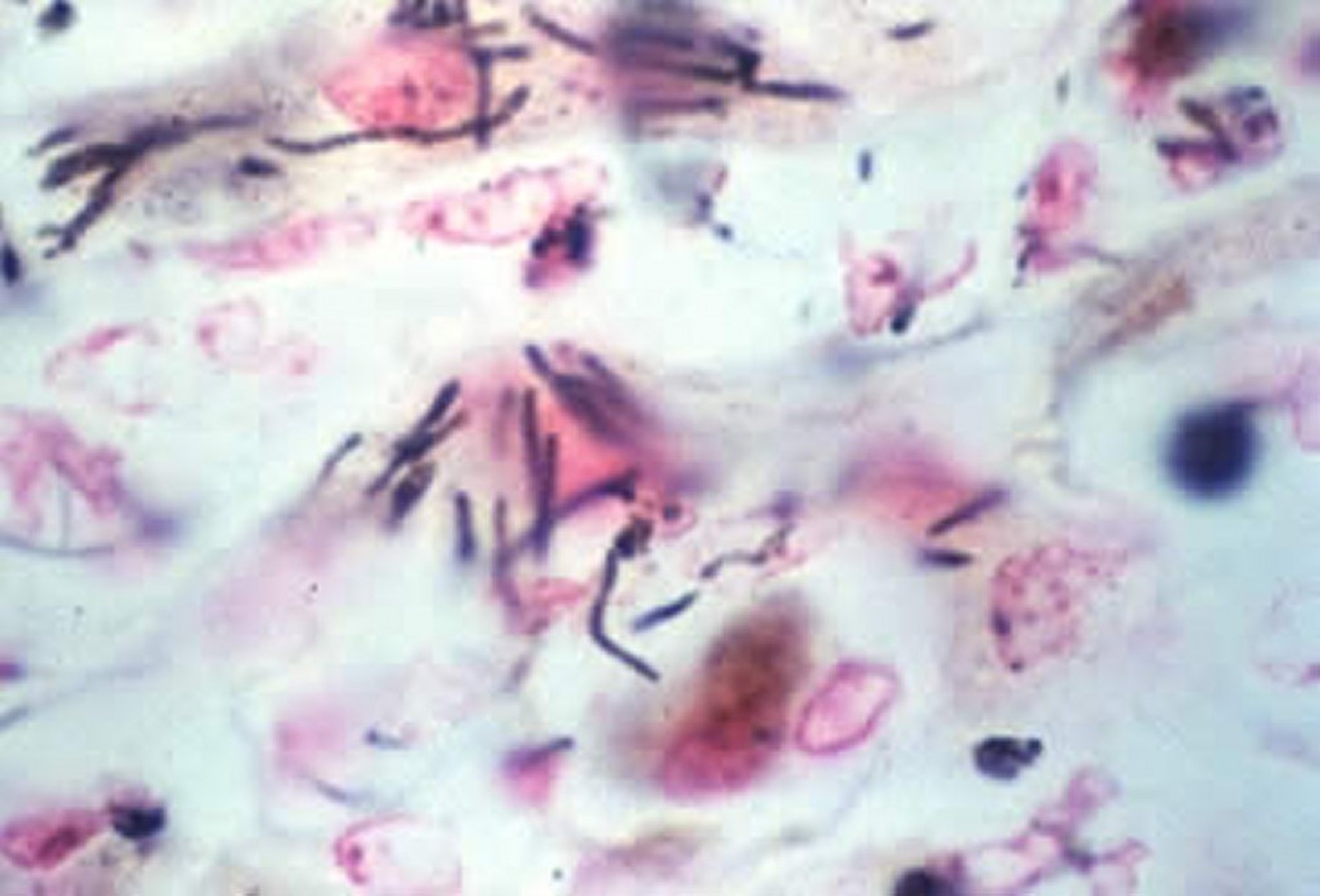
Courtesy of Dr. Francisco Uzal.
Tyzzer disease is caused by Clostridium piliforme, a gram-variable, motile, spore-forming, rod-shaped, flagellated, obligate, intracellular bacterium. It does not grow in cell-free media but can be cultured in the yolk sac of chick embryos or tissue culture cells. The vegetative phase is very labile, but spores may survive in soiled bedding at room temperature for >1 year and are resistant to heating up to 60°C for 30 minutes, or to exposure to 70% ethanol, 3% cresol, 4% chlorhexidine, and 0.037% formaldehyde. However, spores are sensitive to 0.4% peracetic acid, 0.015% sodium hypochlorite, 1% iodophor, and 5% phenol.
C piliforme appears to be common in the environment, but because it is a difficult organism to culture, little is known about the epidemiology, pathogenesis, and immunity of this infection. Most information about the pathogenesis of Tyzzer disease comes from experimental work in laboratory animals; little is known about the pathogenesis of the spontaneous disease in domestic or wild animals. The feces of sick or carrier animals are the primary source of spores that contaminate the environment.
Once the disease is present on a farm, it may be seen sporadically every year. The mode of transmission is believed to be fecal-oral, by ingestion of spores, after which most immunocompetent animals clear the infection within a few weeks. In susceptible individuals, however, C piliforme replicates in the intestinal mucosa, likely the ileum, colon, and cecum, where it results in enterocyte death and, in some cases, associated inflammation. From there, the microorganism is absorbed into the portal circulation, from where it is distributed to the liver and other organs. The bacterium has an affinity for the epithelial and smooth muscle cells of the intestines, hepatocytes, and cardiac myocytes. Stress factors such as capture, overcrowding, shipping, and poor sanitation appear to be predisposing. Sulfonamide administration predisposes rabbits to the disease.
C piliforme infections are often subclinical or asymptomatic; however, they may be severe and lethal in many animal species. There are differences in susceptibility within animal species. B lymphocytes, T lymphocytes, and natural killer cells may play a role in mediating strain susceptibility in laboratory animals. Tyzzer disease is relatively common in horses, which are susceptible to at least two distinct strains, although very little information is available about specific features of disease caused by each of these strains. Some isolates of C piliforme produce toxins; however, others do not. The nature and the role of these toxins in the pathogenesis of infection is unknown, but the toxic isolates are generally more virulent than the nontoxic isolates.
The disease in foals occurs most often between 1 and 6 weeks of age, with most cases occurring between 1 and 2 weeks, and at weaning in rabbits. In some species, Tyzzer disease has been identified concurrently with other diseases, such asfeline infectious peritonitis and feline herpesvirus infection in cats, distemper and mycotic pneumonia in dogs, and cryptosporidial and coronaviral enteritis in calves.
The disease in foals is more common during spring, when nursing mares are exposed to lush, high-protein pastures. The increase in the availability of nutrients from pasture forages and supplemental diets may encourage the overgrowth of C piliforme in the gut of nursing mares. This process predisposes neonatal foals to the disease when they are exposed to massive numbers of the bacterium by ingesting the feces of their dams soon after birth as a mechanism to establish their normal intestinal flora. The immature gut is likely more permeable to pathogens such as C piliforme.
In foals, Tyzzer disease affects primarily the liver, where it induces widespread multifocal necrosis, and foals usually die of acute liver failure. Only young foals up to 6 weeks old develop lesions. In one study in California, all foals with Tyzzer disease had liver lesions, while fewer than one-third of those animals had enteric or myocardial lesions. Older foals become more resistant to the disease as the gut matures. The disease is not recognized in adult horses, but they carry the bacterium in their gut.
In horses, immunologic factors seem to affect the likelihood of disease, because many adults have antibodies against C piliforme and do not develop clinical signs. Tyzzer disease is more common in foals when young, nursing mares are introduced to a farm where the disease is endemic, and it is less common in suckling foals with older mares; this suggests that older mares are immune to the disease and may transfer C piliforme antibodies to their foals in colostrum.
Clinical Findings of Tyzzer Disease in Animals
Tyzzer disease often affects apparently healthy, fast-growing foals. The incubation period in experimentally infected foals is 4–7 days after oral exposure to bacterial spores. Most foals are found in a coma or dead. Clinical signs, if seen, are of short duration, from a few hours to 2 days. Clinical signs are variable but may include depression, anorexia, pyrexia, jaundice, diarrhea, and recumbency. Near death, convulsions and coma occur.
Laboratory animals, including rabbits, may be found dead at the start of an outbreak. As the disease progresses in the colony, animals may show depression, a ruffled coat, and varying extents of watery diarrhea. Clinical signs in other animal species are highly variable.
In foals, the serum enzymes sorbitol dehydrogenase, aspartate aminotransferase, alkaline phosphatase, lactate dehydrogenase, and gamma-glutamyltransferase are increased. Hyperbilirubinemia, leukopenia, hemoconcentration, and terminally profound hypoglycemia may also develop. Clinicopathologic tests are of minimal value in laboratory animals because of quick death.
Lesions
Characteristic gross lesions of Tyzzer disease are seen in the liver, and less frequently in the myocardium and/or intestinal tract. Rarely, lesions in other organs are seen. Throughout the liver of most animal species, many white, gray, or yellowish foci of necrosis, ~2 mm in diameter, may be seen. In addition, hepatomegaly is marked, and the hepatic lymph nodes are edematous and hyperplastic.

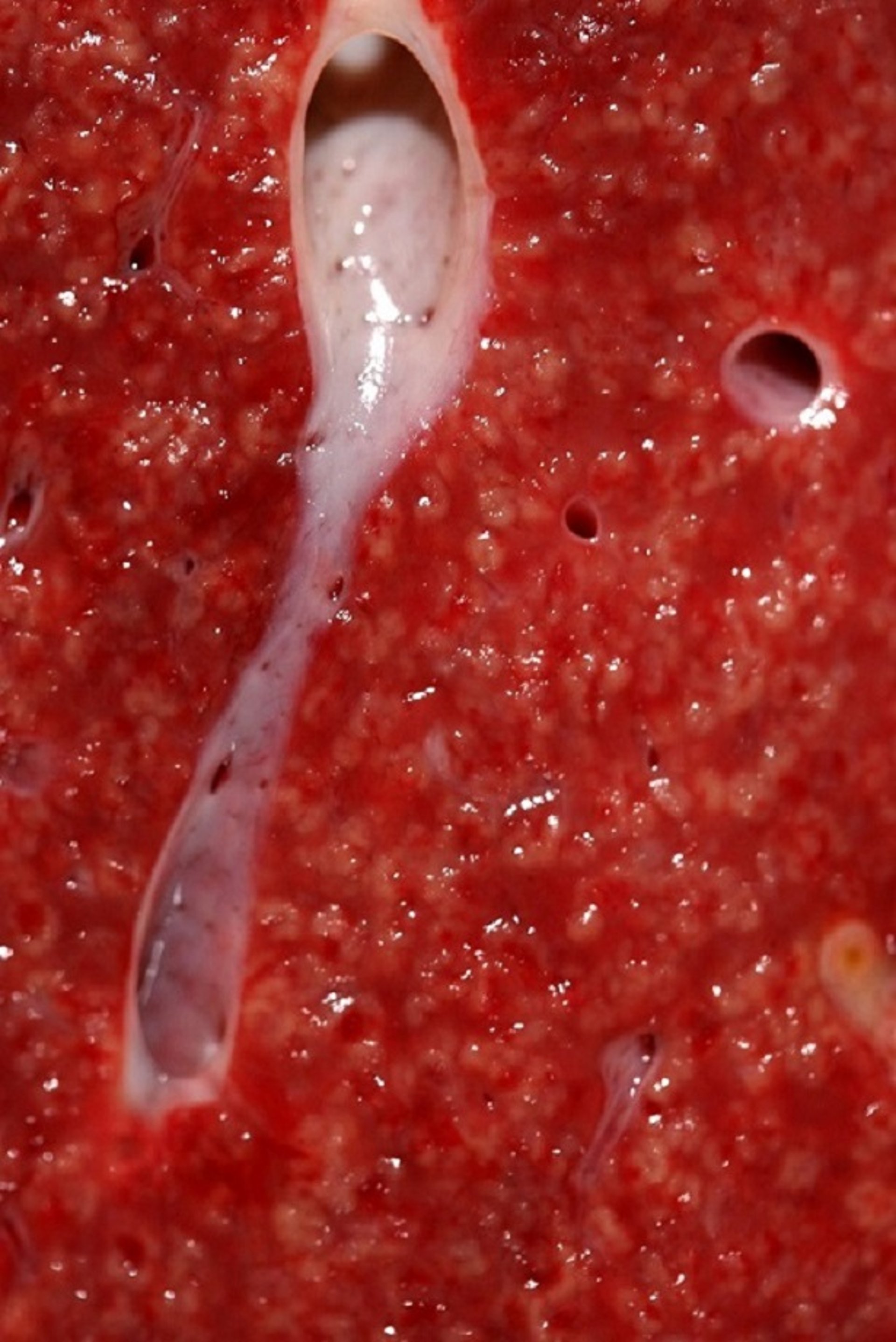
Courtesy of Dr. Francisco Uzal.
Foals with Tyzzer disease always have liver lesions; however, lesions occur much less frequently in the colon and heart. In rabbits, in addition to liver lesions, severe lesions develop in the intestine and heart. The ileum, cecum, and proximal colon are diffusely reddened. Ecchymoses are frequently seen on the serosa of the colon and cecum. Patchy areas of mucosal necrosis and transmural edema are present in these organs. Mesenteric lymph nodes may be enlarged and edematous. White streaks in the myocardium may be present, especially near the apex. Intestinal and heart lesions are generally milder or absent in most animals of other species.

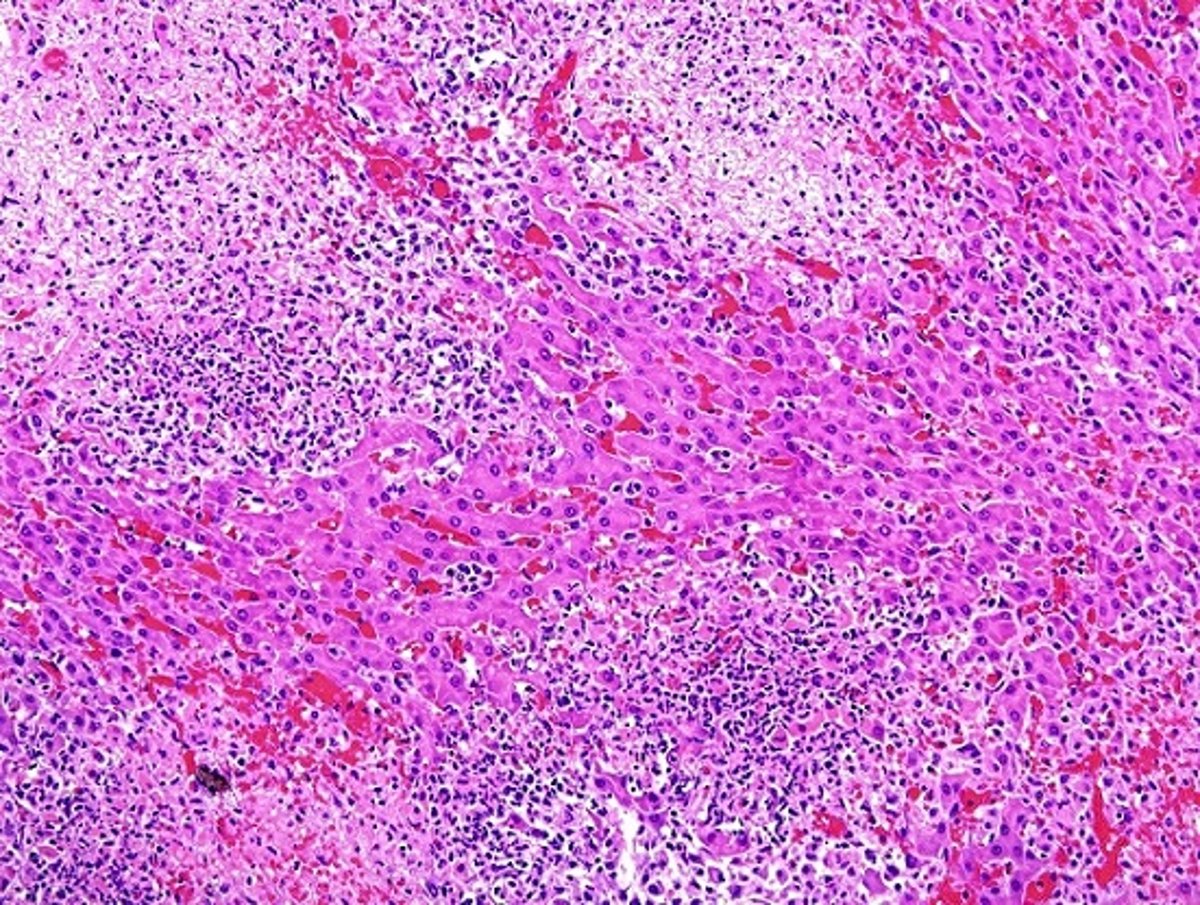
Courtesy of Dr. Francisco Uzal.
Microscopically, in all animal species, numerous multifocal areas of lytic hepatic necrosis can be seen. In foals, the hepatic lesions are more pronounced than in other animals. The necrotic foci tend to coalesce. The hepatocytes in the center of the necrotic foci are replaced by a mixture of cell debris, mononuclear cells, neutrophils, and RBCs. The causative filamentous bacilli are found in a crisscross pattern in the cytoplasm of hepatocytes and rarely in the interstitium, mostly at the periphery of the necrotic foci.

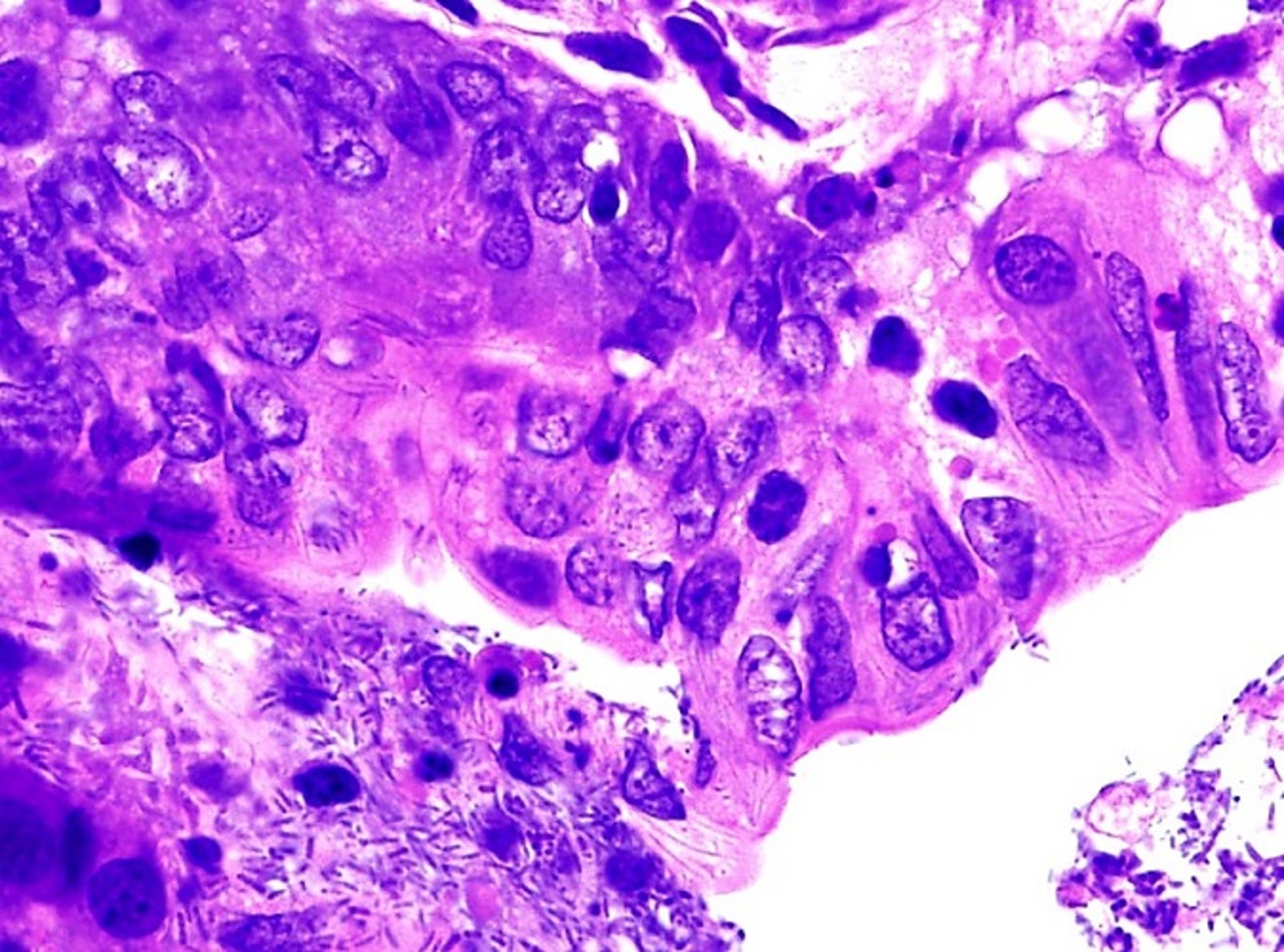
Courtesy of Dr. Francisco Uzal.
In the cecum and colon of rabbits, multifocal mucosal ulceration is observed associated with infiltrates of neutrophils. Bacilli may be found within the enterocytes and the smooth muscle cells of the muscularis mucosa and muscularis externa. When cardiac lesions are present, they consist of foci of myocardial necrosis, with minimal inflammatory cell infiltration.
Lesions in other animal species are variable but tend to resemble those described for foals and/or rabbits, although other systems may also be affected.
Diagnosis of Tyzzer Disease in Animals
On the basis of clinical signs and results of clinical pathologic testing including PCR assay of fecal samples
At post-mortem examination, presumptive diagnosis is based on gross findings, with confirmation based on characteristic histopathologic lesions and presence of C piliformein tissue sections
Clinical signs and results of clinical pathologic testing compatible with liver failure are only suggestive of Tyzzer disease. PCR assay for C piliforme in feces of suspect infected animals adds certainty to a presumptive clinical diagnosis. Because healthy animals can be carriers of C piliforme, however, results of PCR assay must be interpreted with caution.
In dead animals, a presumptive diagnosis of Tyzzer disease is based on gross findings during the necropsy examination, particularly the characteristic multifocal liver necrosis frequently coupled with diffuse icterus. Confirmation of the diagnosis should be based on microscopic lesions, including demonstration of organisms in tissue sections with special stains. C piliforme stains poorly with H&E and Gram stains. With Giemsa or silver (Warthin-Starry, Levaditi, or GMS) stains, the bacilli stain well in the hepatocytes and intestinal epithelium and in smears of infected organs. They are, however, difficult to see in cardiac muscle cells. PCR assay for C piliforme in liver tissue from animals with characteristic gross and microscopic lesions is also diagnostic.
Treatment and Control of Tyzzer Disease in Animals
Supportive care
C piliforme is sensitive to tetracycline and partially sensitive to streptomycin, erythromycin, penicillin, and chlortetracycline. It is resistant to sulfonamides and chloramphenicol. However, there is limited information on the effectiveness of antibiotic treatment of Tyzzer disease.
In neonatal foals, the disease is close to 100% fatal, although occasionally a foal, particularly if older and less severely affected, survives.
A few presumptive cases of Tyzzer disease in foals have been treated successfully with aggressive supportive care, including administration of IV dextrose, sodium bicarbonate, potassium chloride, penicillin, and sulfamethoxazole-trimethoprim.
Because the disease in foals is sporadic and not highly contagious, specific preventive measures are usually not indicated. In areas where spores are present in the environment, many foals may be exposed; however, only a few that are immunosuppressed become acutely affected. On premises where the disease is prevalent, overfeeding of mares, especially with high-protein diets, seemingly predispose neonatal foals. Reducing the nitrogenous dietary compounds, including protein and nitrate in the diet that may induce immunosuppression in neonatal foals, may lessen the incidence of the disease. In general, factors that cause stress and immunosuppression should also be reduced.
When the disease is seen in a colony of laboratory animals, treatment is not recommended because it prolongs the disease and possibly produces carrier animals. It is best to euthanize all animals in the colony, decontaminate the environment, and restock with disease-free animals.
Key Points
Tyzzer disease is a highly fatal disease produced by C piliforme and characterized mainly by liver lesions and failure; lesions in intestine, heart, and other organs are less frequently observed.
Tyzzer disease affects mostly young animals of various species.
A presumptive diagnosis is based on clinical findings, although confirmation should be based on analysis of gross and microscopic lesions and/or PCR assay for C piliforme.
Treatment of Tyzzer disease is supportive and nonspecific; the success rate is usually very low.
For More Information
Fresneda KC, Carvallo Chaigneau FR. Tyzzer’s disease. In: Clostridial Diseases of Animals. Uzal FA, Songer WG, Prescot J, Popof M (eds). Wily and Blackwell; 2016: pp. 281-291.
Sellon DC. Systemic clostridial infections, Tyzzer’s Disease. In: Equine Infectious Diseases. Sellon DC, RW Long (eds). Saunders Elsevier; 2007: pp. 367-372.
Swerczek T. Tyzzer’s disease in foals: retrospective studies from 1969 to 2010. Can Vet J. 2013;54:876-880.




