

Classical swine fever is a highly infectious and often fatal viral disease of swine. Swine infected with classical swine fever virus usually develop fever, hemorrhages, lethargy, yellowish diarrhea, vomiting, and purple skin discoloration of the ears, lower abdomen, and legs. Neurological signs, reproductive failures, and abortion may also be observed. Diagnostic tests for the disease include RT-qPCR assay, virus isolation, immunofluorescence assay, and detection of antibodies by serological tests such as ELISA and virus neutralization. There is no treatment for classical swine fever.
Classical swine fever (CSF) is a highly infectious viral disease; cases should be reported to the World Organisation for Animal Health (WOAH). National animal health authorities report cases to WOAH; hence, suspected cases of CSF are usually reported first to local animal health authorities.
Globally, CSF virus (CSFV) is considered a high-consequence pathogen. There are no treatments for CSFV infection. Instead, vaccination against CSFV with live, attenuated vaccines (LAVs) prevents the disease and is usually performed in regions where CSF is endemic. Countries considered free of the disease do not vaccinate.
Pearls & Pitfalls
|
Infection with highly virulent CSFV isolates generally leads to the death of infected animals, whereas moderate- to low-virulence isolates induce a chronic disease. Suspected cases are confirmed via laboratory testing by officially authorized laboratories.
Etiology of Classical Swine Fever
Classical swine fever is caused by an enveloped RNA virus in the genus Pestivirus of the family Flaviviridae.
CSFV is antigenically related to other pestiviruses, mainly to the bovine viral diarrhea virus (BVDV) of cattle and to the border disease virus (BDV) of sheep. These viruses are highly prevalent in bovine and ovine populations; infection with these viruses in pigs is possible, but uncommon. Transmission of ruminant pestiviruses to pigs usually requires direct contact with cattle, sheep, or goats. In most cases, ruminant pestivirus infections in pigs are rapidly cleared and do not result in clinical signs that are suggestive of CSF. However, infections with either BVDV or BDV induce an antibody response in swine. Therefore, in pigs with suspected CSF, antibody discrimination tests must be done to differentiate CSFV infection from infections caused by ruminant pestiviruses.
CSFV naturally infects members of the Suidae family, ie, domestic and wild pigs. CSFV has only one serotype, but has minor antigenic variability between strains. Strains can be typed for epidemiological purposes by sequencing the entire viral genome or specific regions of the viral genome (ie, 5′UTR, E2, and NS5B regions) combined with phylogenetic analysis.
The virus is moderately fragile and does not persist in the environment or spread long distances by the airborne route. However, it can survive in moist, protein-rich environments such as pork tissues and body fluids, especially if kept cold or frozen. Virus survival times of up to several years have been observed in frozen pork meat. CSFV may also survive for months in chilled or cured cuts.
Epidemiology of Classical Swine Fever
Classical swine fever virus is distributed worldwide (see World Organisation for Animal Health event data). It is considered endemic in certain countries of Central and South America, in the Caribbean basin, and many pork-producing countries in Asia. National pig herds in Australia, New Zealand, Canada, the US, and some European countries are free of the disease. The pig is the primary source of CSFV infection, either via infected live animals or uncooked pork products.
In areas where CSFV is endemic, the primary concern is the spread of disease via the movement of infected animals. Wild boars and species within the Suidae family are susceptible to CSFV infection. In Europe, CSFV is endemic in wild boar populations, which tend to be the source of CSF outbreaks reported among domestic pigs. CSF outbreaks can also be caused by the introduction of the virus into herds via swill feeding.
CSFV is readily inactivated by heat (ie, cooking), emphasizing the importance of heat treatment of swill feed before feeding pigs. Nonetheless, to minimize risks of disease outbreaks, many countries have completely banned swill-feeding practices. The mechanical transmission of CSFV by vehicles, equipment, and personnel traveling between pig farms is also implicated in disease spread. The persistence of CSFV within herds for long periods has been observed.
Infections of sows during pregnancy with minimally to moderately virulent strains of CSFV may lead to in utero infections of fetuses. These infections lead to litters born persistently infected with CSFV, which are carriers of the virus and a source of new infections. Persistently infected carrier pigs usually do not show clinical signs but constantly shed CSFV into the environment. Therefore, it is imperative to consider CSFV infections while investigating herds presenting with unexplained reproductive failures or clinical signs in piglets, such as tremors or other congenital abnormalities.
The widespread use of CSFV vaccines in China is believed to have led to CSFV escape variants, which are not fully neutralized by the vaccine-derived immune response. CSFV isolates from genotype 2.1 are neutralized to a lesser extent by sera from pigs immunized with LAV C strain than isolates of genotypes 2.2 and 2.3. The connection of these findings to potential vaccine failures is yet to be determined.
Clinical Findings and Lesions of Classical Swine Fever
Classical swine fever is characterized by fever, hemorrhages, ataxia, and purple skin discoloration; however, the clinical presentation varies depending on host characteristics and the particular virus strain causing the infection. The principal lesion produced by CSFV infection is a generalized vasculitis, clinically manifested as hemorrhages and cyanosis of the skin, notably at the ears, lower abdomen, and extremities (see ). There may also be a generalized erythema of the skin. Vasculitis in the CNS leads to incoordination (ie, staggering gait) or even convulsions. Histologically, nonsuppurative encephalitis with a characteristic vascular cuffing is commonly observed.

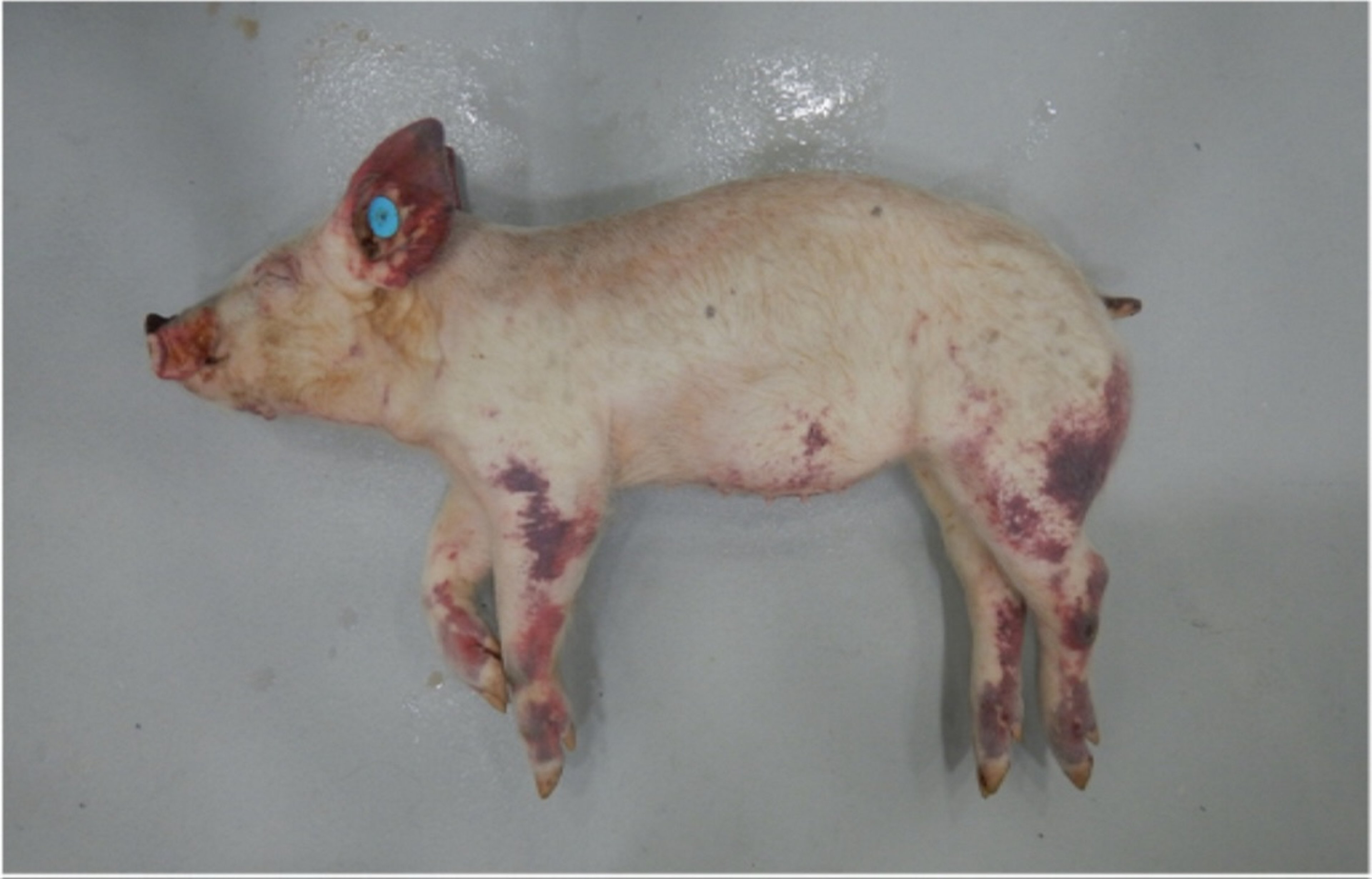
Hemorrhagic skin discoloration observed in the muzzle, pinna, ventral abdominal wall, and limbs of a piglet infected with CSFV strain Diepholz, a subgenotype 2.3 virus, isolated in Germany, 1994.
Courtesy of Dr. Aruna Ambagala and Dr. Carissa Embury-Hyatt.
CSF occurs in several forms, including acute, chronic, and subclinical.
Acute forms of CSF, associated with highly virulent CSFV strains, are characterized by an incubation period of 3–7 days, with death occurring within 10 days after infection. Fever > 41°C (105.8°F) is usually observed and persists until the terminal stages of the disease, when body temperature drops and becomes subnormal. Constipation, followed by diarrhea and vomiting, is common. At necropsy, the principal findings are widespread petechial and ecchymotic hemorrhages, especially in lymph nodes (eg, mandibular, retropharyngeal, gastrohepatic), tonsils, larynx, kidneys, spleen, urinary bladder, and ileum (see , , and hemorrhage photographs).


Petechial hemorrhages observed in the stomach mucosa of swine infected with CSFV strain Diepholz, a subgenotype 2.3 virus, isolated in Germany, 1994.
Courtesy of Dr. Aruna Ambagala and Dr. Carissa Embury-Hyatt.

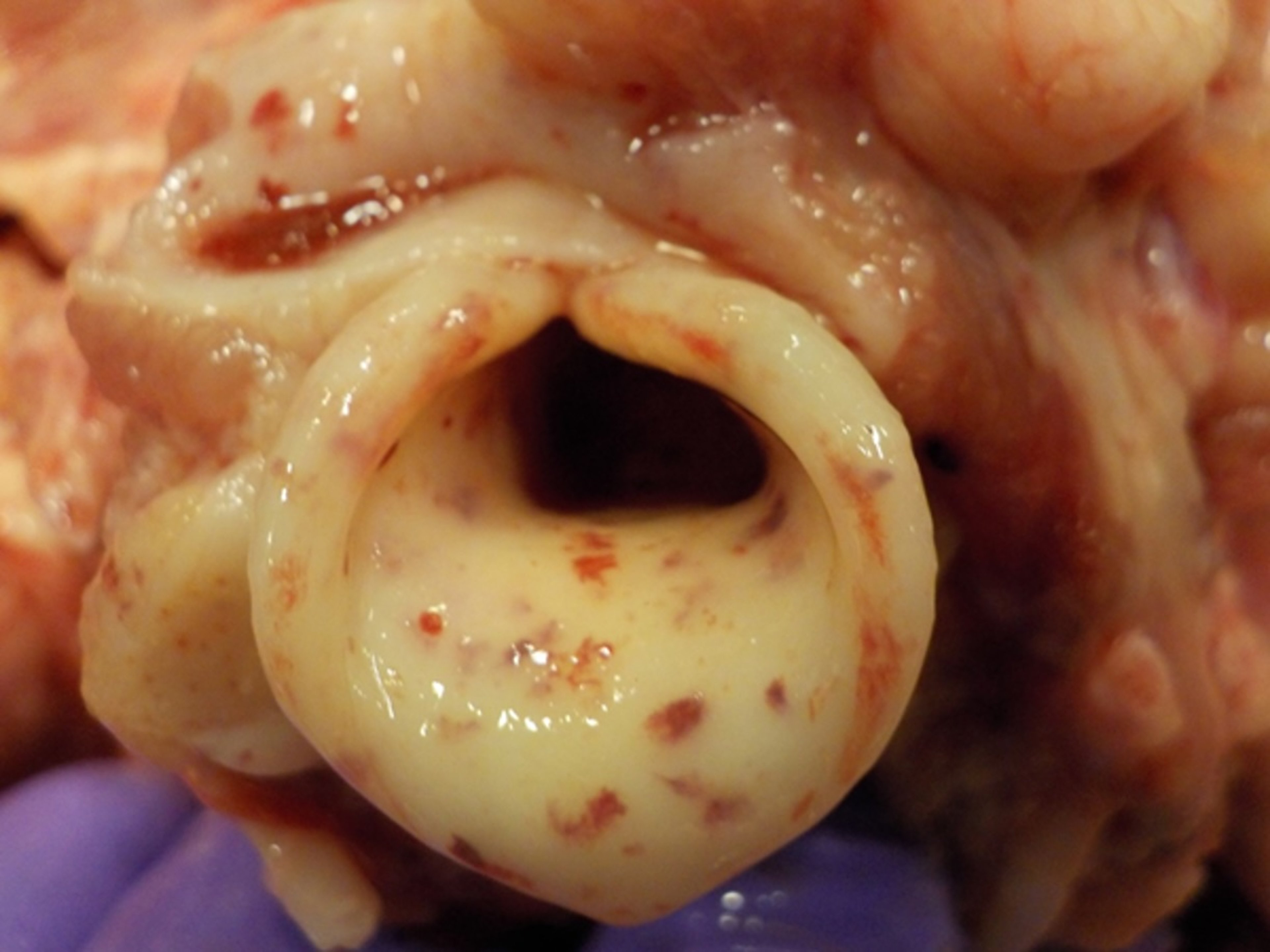
Petechial hemorrhages on the epiglottis of a pig infected with CSFV strain Diepholz, a subgenotype 2.3 virus, isolated in Germany, 1994.
Courtesy of Dr. Aruna Ambagala and Dr. Carissa Embury-Hyatt.

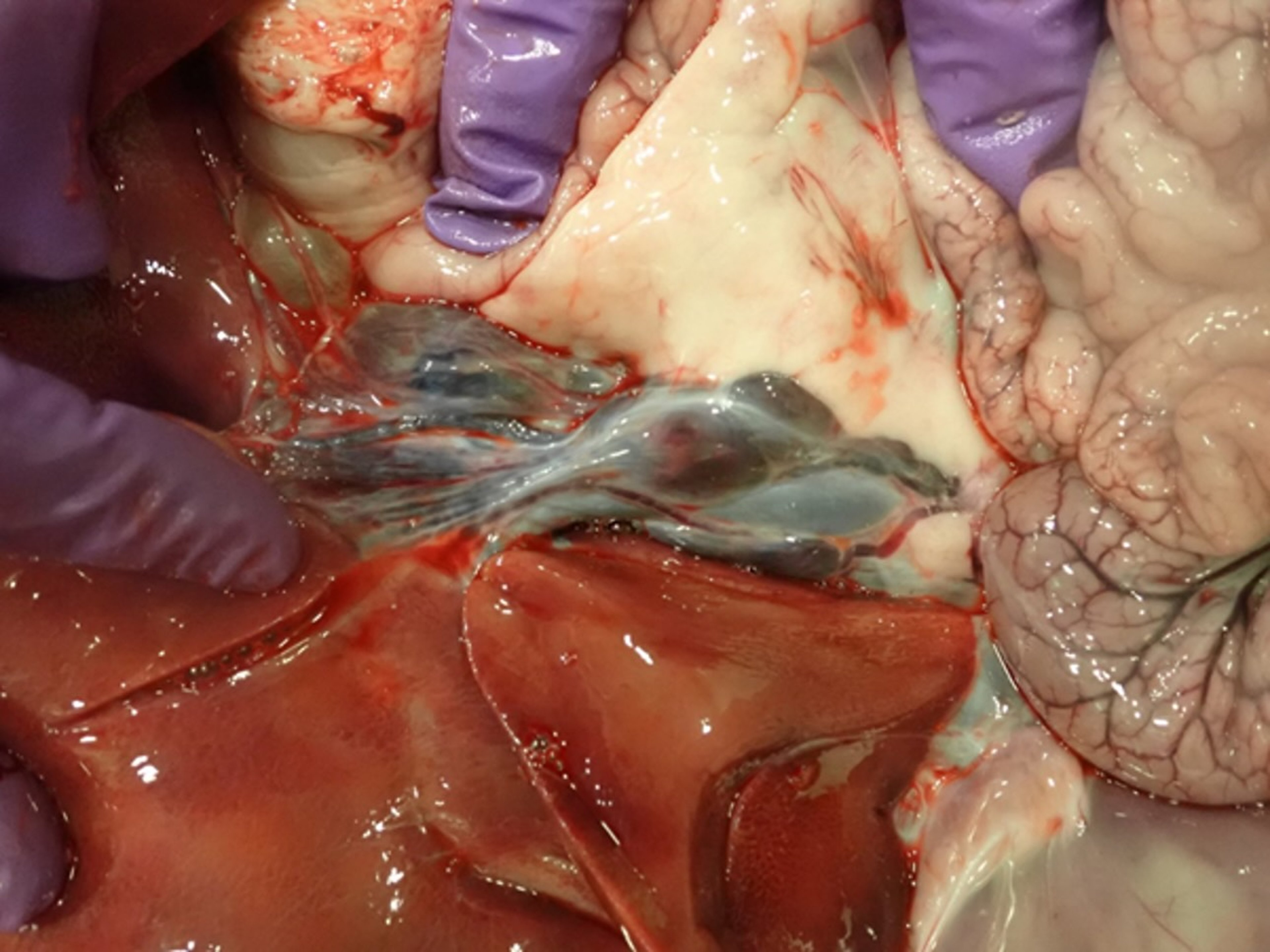
Hemorrhagic gastrohepatic lymph node of a pig infected with CSFV strain Diepholz, a subgenotype 2.3 virus, isolated in Germany, 1994.
Courtesy of Dr. Aruna Ambagala and Dr. Carissa Embury-Hyatt.
Infarction may be observed, particularly in the periphery of the spleen. Macroscopically, in addition to the lesions described above, "button" ulcers may develop in the intestine, particularly near the ileocecal junction. Histologically, atrophy of the thymus and depletion of lymphoid follicles in lymph nodes are present. In the subacute form, death generally happens within 20–30 days after infection; death may occur much later in the chronic condition.
Chronic forms of the disease are also characterized by high fever, staggering gait, cough, diarrhea, purple discoloration of the skin, and death. In chronic forms of CSF, after an initial acute febrile phase, infected animals may show an apparent recovery but then relapse, with anorexia, depression, fever, and progressive loss of condition (ie, marked weight loss). Subacute and chronic forms of the disease are associated with CSFV strains of moderate to low virulence, respectively.
Low-virulence strains can be difficult to detect and are the cause of subclinical presentations; the only clinical expression may be the poor reproductive performance of sows and the birth of piglets with neurological defects (eg, congenital tremor). Advice on pig samples that can be used for detecting CSFV should be sought from the receiving laboratory. Tonsils, lymph nodes (mandibular, retropharyngeal, gastrohepatic, and mesenteric), spleen, kidney, and ileum are suitable for detecting the virus.
Whole blood collected with EDTA as an anticoagulant can be used for virus isolation or virus detection, particularly during the viremic phase of the infection. Clotted blood samples (serum) are taken when serological tests for detecting CSFV antibodies are pursued. Nasal swabs and tonsil scrapings are commonly collected clinical samples that detect the virus (ie, viral RNA). The oronasal route sheds substantial amounts of CSFV from infected animals into the environment, particularly early during infection.
Diagnosis of Classical Swine Fever
RT-PCR
Antigen detection by direct immunofluorescence
Diagnostic tests for detecting CSFV: The diagnosis of CSF is based on clinical signs, serological testing, antigen detection in tissues, virus isolation, and detection of CSFV RNA. Because clinical signs in CSFV-infected pigs are also observed in other diseases of swine, laboratory confirmation of the disease is always required.
Clinically, the differential diagnosis varies based on the course of CSF. African swine fever is the main differential because this disease has a similar clinical presentation (see table ). Hemorrhagic lesions, systemic diseases of pigs, and reproductive failures caused by other infectious and noninfectious agents should also be considered.
Pearls & Pitfalls
|
Serological testing: Antibodies against CSFV are detected later during the infection (2–3 weeks postinfection). ELISA and virus neutralization assays are commonly used to detect CSFV-specific antibodies. It is important to consider that BVDV and BDV can infect pigs and cross-react in ELISA. The presence of antibodies against ruminant pestiviruses in pigs may hamper the serological diagnosis of CSF. Both BVDV- and BDV-specific antibodies are sporadically detected in pig populations. This method is preferred for CSF surveillance.
Antigen detection: CSFV antigen can be detected using direct immunofluorescence on frozen tissue sections, particularly in tonsil samples, using specific antibodies. Antigen detection can also be done using ELISA; however, this assay has low sensitivity and is only useful for screening for CSFV at the herd level.
Detection of CSFV RNA: RT-PCR and RT-qPCR assays are broadly used to rapidly detect CSFV RNA in tissues, blood, serum, or oronasal fluids. The precise assay can differentiate CSFV from bovine viral diarrhea virus and BDV. Standardized RT-PCR methods can be scaled up to screen large numbers of samples, giving rapid results while retaining high sensitivity. This method is particularly useful for screening herds during an outbreak of CSF.
Virus isolation: CSFV is isolated in cell cultures inoculated with tissue suspensions, WBCs, or oronasal fluids of suspected CSF cases. Although accurate, virus isolation can take days to weeks. Instead, preclinical and field-deployable rapid detection tests are widely used.
Classical Swine Fever Differential Diagnoses
Classical Swine Fever Differential Diagnoses | Clinical Similarities |
|---|---|
Parvovirus | Hemorrhages, reproductive failures |
Hemorrhages, reproductive failures | |
Hemorrhages, reproductive failures | |
Hemorrhages | |
Anticoagulant poisoning (eg, warfarin) | Hemorrhages |
Hemolytic disease of the newborn | Hemorrhages |
Postweaning multisystemic wasting syndrome | Hemorrhages |
Salt poisoning | Hemorrhages |
Thrombocytopenic purpura | Hemorrhages |
Systemic disease | |
Eperythrozoonosis | Systemic disease |
Systemic disease | |
Haemophilus suis | Systemic disease |
Systemic disease | |
Pestivirus infections | Systemic disease |
Systemic disease |
Control of Classical Swine Fever
Elimination of exposed herds
Vaccination
Commercial CSF live, attenuated vaccines (LAVs) are safe and highly efficient, inducing protection within the first week after vaccination. These LAVs, originally C strain, have been used worldwide for over 50 years with an extraordinary safety record. Oral vaccination of wild boars has been used successfully within the European Union using a live, attenuated vaccine delivered via baits. Oral vaccination has been a critical strategy to control CSF, mainly where parenteral vaccine delivery is not feasible.
In the 1990s, subunit CSFV vaccines containing only the major viral surface glycoprotein of the virus were licensed. Although these vaccines allow the differentiation of infected from vaccinated animals (DIVA capabilities), they lack the efficiency of live, attenuated vaccines, requiring 3–4 weeks or revaccination to induce solid protection. New generations of CSFV marker vaccines have been developed to make emergency vaccination compatible with the control of CSF.
In 2014, the European Medicines Agency licensed a live, attenuated marker vaccine (1). The vaccine is based on a pestivirus chimera that carries the E2 glycoprotein from a CSFV isolate in a bovine viral diarrhea virus type-1 backbone. Pigs vaccinated with genetically modified viruses generate an antibody response that can be distinguished from the response elicited against CSFV isolates. The protective efficacy induced by these viruses is comparable to the immunity induced by a first-generation LAV.
The application of a marker vaccine is possible if tests such as ELISA can distinguish between antibodies produced in response to natural infection and those created by vaccination. The DIVA principle is based on detecting CSFV-specific antibodies that develop in the host only with CSFV infection but not after vaccination with a marker vaccine. These assays have been designed as necessary companion tests for development and licensing of CSF marker vaccines. Culling of exposed herds is used to prevent disease expansion during outbreaks in nonendemic countries. Highly effective live, attenuated vaccines are widely used in endemic areas to control CSF spread.
Classical swine fever is a reportable disease. Control is usually tightly regulated by local laws that establish strict sanitary measures. No treatment is available.
Countries regain their CSF-free status (no antibodies or virus detected in animals) after establishing that CSFV is no longer present in the national pig herd. Countries free of CSF forbid the use of prophylactic vaccination and exert strict control over the movement of animals, including domestic pigs. In these countries, herds affected by an outbreak of CSF are quarantined, and exposed animals are culled. Sometimes pigs are preemptively culled within established quarantine zones. Emergency ring vaccination around the outbreak can prevent the further spread of CSFV.
In countries where the virus is endemic, prophylactic vaccination is used. If systematically applied, vaccination may lead to the elimination of CSFV from swine herds.
Key Points
Classical swine fever is a highly infectious, often fatal, notifiable viral disease of swine.
CSF is endemic in many countries in Central and South America, the Caribbean, and Asia.
No treatment for CSF is available.
Vaccination with highly effective vaccines is used where the disease is endemic to prevent CSFV expansion.
Vaccination is not used in countries free of the disease. Instead, rapid confirmation of suspected cases, followed by quarantine and depopulation of exposed herds, is applied.
For More Information
Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2023. Twelfth edition. WOAH.
Classical Swine Fever. Center for Food Security and Public Health.
References
Blome S, Wernike K, Reimann I, König P, Moß C, Beer M. A decade of research into classical swine fever marker vaccine CP7_E2alf (Suvaxyn® CSF Marker): a review of vaccine properties. Vet Res. 2017;48(1):51. doi:10.1186/s13567-017-0457-y


