




Adenovirus of Marine Mammals
Adenovirus has been isolated from a sei whale (Balaenoptera borealis), beluga whale, and bowhead whales (Balaena mysticetus). A novel otarine adenovirus 1 has been isolated from California sea lions affected with hepatic disease, and adenovirus particles have been reported in walrus and fur seals. Fatal disease associated with adenovirus infection has been reported in California sea lions, South African fur seals, and South American sea lions. No disease was noted in the cetaceans.
The most prominent histologic lesion in all cases was hepatic necrosis. Massive coagulation necrosis without apparent zonal distribution was seen in some animals. Basophilic intranuclear inclusions in hepatocytes or granular amphophilic intranuclear inclusions in Kupffer cells were seen. No evidence of adenovirus was detected in the lungs. Adenovirus from California sea lions is not known to cause disease in humans.
Caliciviruses of Marine Mammals
Caliciviruses have been isolated from otariid seals, walrus, Atlantic bottlenose dolphins, and opaleye fish (Girella nigricans). The marine caliciviruses appear to be serotypes of vesicular exanthema of swine virus (VESV). Several species of mysticete cetaceans have antibodies to different serotypes of VESV. By 4 months of age, most California sea lions have neutralizing antibodies to one or more of the serotypes. Opaleye fish are probably responsible for the endemic status of caliciviruses in marine mammals that inhabit the coastal waters of California. To date, infections have not been diagnosed in marine mammals in the Atlantic Ocean.

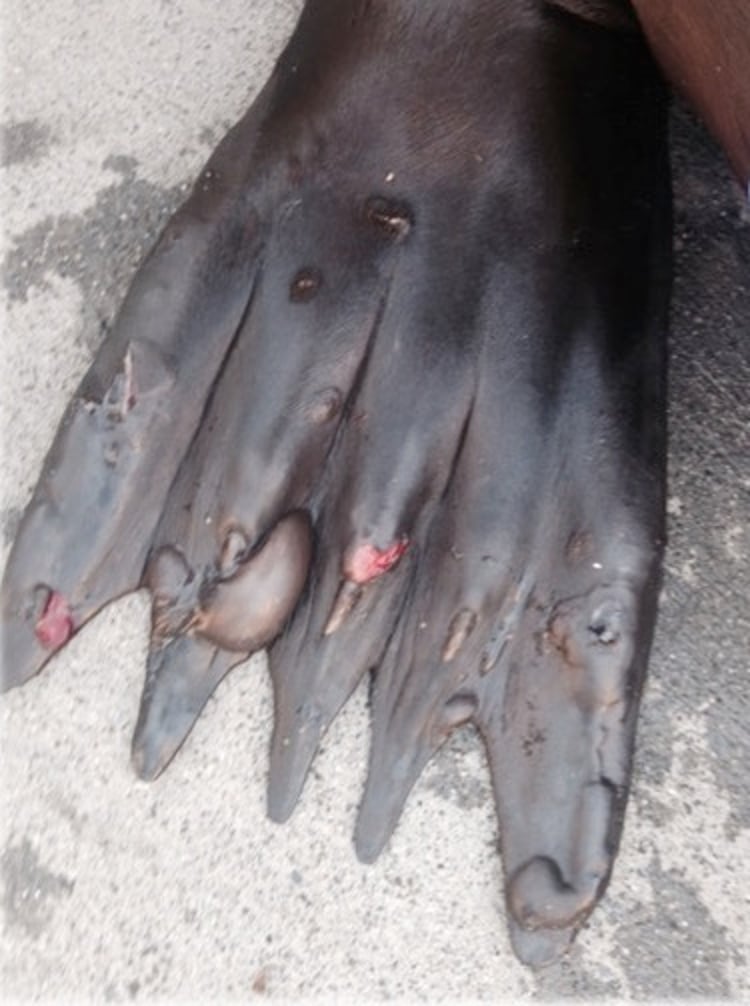
Courtesy of Dr. Cara Field.
The most consistent lesion in marine mammals is skin vesicles. In pinnipeds, the vesicles are prevalent on the dorsal and ventral surfaces of the fore and hind flippers around the nail beds and flipper tips. In dolphins, vesicular lesions have been seen in association with tattoo-like skin lesions and old scars. Vesicles are 1 mm to 3 cm in diameter. They usually erode and leave shallow, fast-healing ulcers, but occasionally vesicles regress and leave plaque-like lesions.
Treatment is supportive only; skin lesions usually resolve without treatment. Infection may cause premature parturition in pinnipeds. Affected pups may have interstitial pneumonitis and encephalitis and fail to thrive, but infection is usually self-limiting.
Inoculation of marine caliciviruses into pigs causes vesicular lesions identical to those seen in vesicular exanthema. In humans, heavy exposure to marine caliciviruses can result in neutralizing antibodies. Localized lesions in an accidental laboratory exposure as well as isolation of calicivirus from a clinically ill primate indicate that these viruses should be handled carefully.
Herpesvirus of Marine Mammals
Herpesviruses isolated from pinnipeds include phocid herpesvirus (PhHV) 1, an alpha herpesvirus, as well as four gamma herpesviruses: PhHV-2, Hawaiian monk seal herpesvirus, otariid herpesvirus 1 (OtHV-1), and Northern elephant seal herpesvirus. Herpesviruses have been identified in a wide range of cetaceans, some associated with encephalitis and others with various skin lesions. Otariid herpesvirus 1 is associated with neoplasia in California sea lions.
Young harbor seals from Atlantic waters infected with PhHV-1 develop nasal discharge, inflammation of the oral mucosa, vomiting, diarrhea, and fever, followed by coughing, pneumonia, anorexia, and lethargy that can result in death in 1–6 days. Morbidity can approach 100% in stressed seals in crowded conditions; mortality is ~50%. The incubation period appears to be 10–14 days. Pacific harbor seals with PhHV-1 tend to develop signs related to adrenal and hepatic dysfunction as well as ocular and oral ulcers, and disease can also be severe.
Phocid herpesvirus 2 has been associated with recurring circumscribed areas of alopecia ~0.5 cm in diameter in gray seals. Herpetic lesions in beluga whales are generally circular, as much as 2 cm in diameter, and may appear slightly depressed with a target appearance or be raised and proliferative. The centers of some lesions are necrotic or may contain verrucous growths. Systemic infections have not been documented in cetaceans.
Diagnosis is often made at necropsy or by clinical signs and observation of characteristic intranuclear inclusion bodies in biopsies of early skin lesions. In seals, interstitial pneumonia caused by herpesvirus must be distinguished from bronchial pneumonia caused by influenza virus.
In systemic herpesvirus infection, therapy is supportive. In a documented epidemic, orally administered acyclovir did not eliminate the infection but appeared to considerably shorten clinical signs in primary infections. Vaccination with 1 mL of trivalent poliovirus vaccine to control recrudescence of suspected herpesvirus lesions has been used with some success. Although it reduced the severity of recrudescence in seals, there is a potential public health risk because live poliovirus may be shed after vaccination. Stress and immunosuppression are associated with recrudescence of latent infections. There is no evidence that the herpesviruses of pinnipeds or cetaceans are zoonotic.
Influenza Virus of Marine Mammals
Four influenza A viruses have been isolated from harbor seals and two other subtypes from a stranded pilot whale. Influenza A subtype H1N1 has been isolated from healthy northern elephant seals, and harbor seals and sea lions have been positive by results of serologic testing. Serologic evidence of exposure to influenza virus exists for a wide range of marine mammals and infection is probably common. Only nonspecific clinical signs were reported in the stranded pilot whale, which had difficulty maneuvering, was emaciated, and was sloughing skin. Disease due to influenza virus in seals is better characterized. Even well-fed captive animals become weak, incoordinated, and dyspneic. Swollen necks due to fascial trapping of air escaping through the thoracic inlet is reported. Occasionally, white or bloody nasal discharge is evident. The incubation period during epidemics in harbor seals is ≤3 days. Many factors probably contribute to the explosive nature of the reported epidemics. High population densities and unseasonably warm temperatures contribute to high mortality.
Diagnosis can be confirmed by means of virus isolation or PCR assay. In harbor seals, influenza pneumonia is characterized by necrotizing bronchitis and bronchiolitis and hemorrhagic alveolitis. In the pilot whale, the lungs were hemorrhagic and a hilar node was greatly enlarged. For differential diagnosis, see Herpesvirus.
The virulence of epidemics has precluded attempts at intensive supportive care. Humans whose eyes were contaminated while doing necropsies, or by being sneezed on by affected seals, have developed keratoconjunctivitis within 2–3 days, and identical virus has been recovered. All affected humans recovered completely within 7 days without developing any antibody titers, which suggests that the reaction is local, as occurs with Newcastle disease virus.
Morbilliviruses of Marine Mammals
Morbillivirus has been associated with mass mortality events in many odontocete and phocid populations and appears to be highly contagious among marine mammals. Phocid seals are susceptible to canine distemper virus (CDV) and to a closely related but distinct morbillivirus, phocine distemper virus (PDV).
Generally, young seals are affected and develop letthargy, anorexia, crusting conjunctivitis, nasal discharge, and dyspnea as well as abnormal neurologic signs. Pneumonia and CNS disease develop, and mortality can be high in previously unexposed animals. Outbreaks of PDV in wild harbor seals have been extensive in the North Sea, and CDV has caused death in Caspian and Baikal seals. Seals vaccinated with CDV vaccine have been rendered immune to challenge with the virus (suspension of organ material) obtained from dead wild seals. Harp seals are thought to be reservoirs for PDV in wild populations.
Cetacean morbillivirus (CMV) is closely related to rinderpest and peste des petits ruminants. Multiple strains of CMV exist, and these viruses have been implicated in the deaths of harbour porpoises and common dolphins off the coast of the UK, striped dolphins in the Mediterranean, and bottlenose dolphins in the western Atlantic and Gulf of Mexico. Pilot whale calves and white-beaked dolphins (Laegenorhynchus albirostris) have also been affected by CMV. Pilot whales are thought to be reservoirs of CMV.
Evidence of morbillivirus exposure has been documented in sea otters, particularly CDV, but infection with PDV may also have occurred. Polar bears have serologic evidence of morbillivirus exposure, although deaths have not been reported. Morbillivirus had been documented in the Florida manatee but no deaths have been documented in this or other sirenians.
Treatment is supportive. Mortality in naive populations is high, often due to secondary infections facilitated by the immunosuppressive impact of active morbillivirus infection. Vaccination with a subunit vaccine is practiced in European rescue centers and appears to be protective. A vaccination program in Hawaiian monk seals has been implemented using a recombinant canarypox-vectored canine distemper vaccine and seals have successfully seroconverted.
Poxvirus of Marine Mammals
Poxvirus has been identified morphologically in skin lesions of both captive and free-ranging pinnipeds and cetaceans of a wide range of species. Lesions in California sea lions, harbor seals, and gray seals are probably due to parapoxviruses; lesions in South American sea lions (Otaria byronia) and northern fur seals are probably not. An orthopox virus has been isolated from pox-like lesions on a gray seal. Poxvirus has also been associated with skin lesions in Atlantic bottlenose dolphins, Atlantic white-sided dolphins, killer whales, dusky dolphins, long-beaked common dolphins (Delphinus capensis), Hector’s dolphins (Cephalorhynchus hectori), and Burmeister’s porpoises (Phocoena spinipinnis), among others.


Courtesy of Dr. Cara Field.
Outbreaks typically occur in postweanling pinnipeds recently introduced into captivity. The incubation period is 3–5 weeks. A break in the epithelial surface may be required to start an infection. Lesions can recur. Small, cutaneous, raised nodules (0.5–1 cm in diameter) occur on the head, neck, and flippers of affected pinnipeds. These may increase to 1.5–3 cm in diameter during the first week and may ulcerate or develop satellite lesions during the second week. After the fourth week, lesions begin to regress, although nodules are reported to persist as long as 15–18 weeks. Areas of alopecia and scar tissue may remain after resolution.
Cutaneous poxvirus infections in cetaceans can develop on any part of the body, but lesions are most common on the head, pectoral flippers, dorsal fin, and tail fluke. They range from ring or pinhole lesions to black, punctiform, stippled patterns (tattoo-like skin lesions). Ring or pinhole lesions appear as solitary, 0.5–3 cm, round or elliptical blemishes, which sometimes coalesce. They are usually light gray and may have a dark gray border, although the reverse color pattern is also seen. Lesions may persist for months or years without apparent ill effects.
Major differential diagnoses include cutaneous streptothricosis and calicivirus infection. Diagnosis is based on the presence of eosinophilic, intracytoplasmic inclusion bodies in lesion biopsies and is confirmed by identification of typical poxvirus particles by electron microscopy.
Poxviruses of marine mammals do not appear to cause systemic infections. Although animals with cutaneous poxvirus lesions have died, other factors were responsible. Therapy to control secondary bacterial infections is indicated only when skin lesions suppurate. The parapoxviruses of pinnipeds can cause isolated lesions on the hands of humans not wearing gloves during contact with infected animals.
Miscellaneous Viral Diseases of Marine Mammals
A ringed seal (Phoca hispida) in Norway was diagnosed with rabies, which was confirmed by immunofluorescent examination of the brain. At the time, there was an epidemic of rabies in foxes in the area. Rabies has also been reported in a polar bear. Other rhabdoviruses isolated from cetaceans, which are not recognized by antisera to representatives of the Lyssavirus, Ephemerovirus, or Vesiculovirus genera, may be related to rhabdoviruses of fish.
Papillomavirus infections have been reported in a wide range of cetaceans, including narwals (Monodon monoceros) and several species of mysticete whale. Lesions are typical of those found in terrestrial species. No therapy is known. Lesions are usually self-limiting.
Hepadnavirus infection with a hepatitis B–like agent has been documented in a Pacific white-sided dolphin with a long history of recurrent illness in captivity. No evidence of zoonotic transmission was identified.
Endogenous gamma retroviruses have been identified in the genome of killer whale, bottlenose dolphin, Atlantic white-sided dolphin, long-finned pilot whale, striped dolphin, short-beaked common dolphin, Risso's dolphin, harbour porpoise, and white-beaked dolphin. The only retrovirus cultured and characterized to date in a marine mammal was a spumavirus isolated from recurring skin lesions in a California sea lion that subsequently died of Pasteurella pneumonia complicated with herpesvirus.
Coronavirus infections have been associated with pneumonia epizootics in wild Pacific harbor seals. A coronavirus has also been identified in a beluga whale.
An enterovirus of unknown pathogenicity isolated from a rectal swab of a California gray whale has been reclassified as a calicivirus. Antibodies, unassociated with disease, against human influenza virus (after challenge) and poliomyelitis virus were found in bottlenose dolphins. Enterovirus has been found associated with tongue lesions in bottlenose dolphins.
Severe enteritis and vomiting that rapidly led to death in a captive beluga whale were suggestive of parvovirus enteritis, but no virus was isolated.







