



Avian leukosis is a neoplastic disease of poultry characterized by tumors of hemopoietic tissues and sarcomas, including lymphoid leukosis, myeloid leukosis, and erythroid leukosis, induced by a related group of avian leukosis/sarcoma viruses. Disease usually occurs in chickens aged 16 weeks or older. Standard criteria used for diagnosis include history, clinical signs, gross necropsy, and histological examination. There are no effective treatments or vaccines; therefore, eradication of the virus from breeding flocks is the most effective control method.
Under natural conditions, lymphoid leukosis has been the most common form of the leukosis/sarcoma group of diseases observed in chicken flocks, although in the 1990s myeloid leukosis become prevalent in meat-type chickens.
The International Committee on Taxonomy of Viruses has designated avian leukosis virus as the type species within the Alpharetrovirus genus of the family Retroviridae. Members of this RNA group of viruses have similar physical and molecular characteristics and share a common group-specific antigen. Detection of the major antigen (p27) present in the core of leukosis/sarcoma viruses forms the basis of several diagnostic tests.
Lymphoid leukosis occurs naturally only in chickens. Tumor mortality commonly accounts for ~1%–2% of birds, with occasional losses of ≥ 20%.
Subclinical infection, to which most flocks are subject, decreases several important performance traits, including egg production and quality. The frequency of infection has been decreased substantially in the primary breeding stocks of several commercial poultry breeding companies, particularly egg-type breeders, and infection has become infrequent or absent in certain commercial flocks. The frequency of lymphoid leukosis tumors even in heavily infected flocks is typically low (< 4%), and disease is often inapparent. However, as much as 1.5% excess mortality per week has been reported in commercial broiler-breeder flocks naturally infected with subgroup J avian leukosis virus.
Etiology of Avian Leukosis in Poultry
Lymphoid leukosis is caused by certain members of the leukosis/sarcoma group of avian retroviruses. Isolates that can induce lymphoid leukosis in chickens are commonly called avian leukosis viruses and are divided into subgroups A, B, C, D, and J, on the basis of differences in their viral envelope glycoproteins, which determine antigenicity, viral interference patterns with members of the same and different subgroups, and host range.
Subgroups A and B are most prevalent in Western countries. Since the initial isolation of subgroup J avian leukosis virus in England, the virus has been isolated from broiler-breeder stocks that experience myeloid neoplasms (myelocytoma) in many other countries. A sixth subgroup (E) designates nononcogenic endogenous viruses produced by viral genes integrated into the host cell DNA. All field strains of avian leukosis virus are oncogenic, although some differences in oncogenicity and replicative ability have been recognized. Recombination between two different subgroups of avian leukosis virus can occur in field conditions and cause economic losses. A new subgroup K has been reported from China; however, its status as a new subgroup remains to be confirmed because it still uses the same host cell receptor as subgroup A (1).
Pathogenesis of Avian Leukosis in Poultry
Lymphoid leukosis is a clonal malignancy of the bursal-dependent lymphoid system. Transformation invariably occurs in the intact cloacal bursa, often as early as 4–8 weeks after infection. Tumors are often not detectable until chickens are ~14 weeks old. Death rarely occurs before 14 weeks and is more frequent around the time of sexual maturity.
The disease can be prevented, even in chickens up to 5 months old, by treatments that destroy the cloacal bursa. The tumors are composed almost entirely of B lymphocytes that, in many instances, have IgM on their surfaces. No antitumor immune response has been recognized. Antibodies are readily induced after infection, except when tolerance occurs.
The induction of lymphoid leukosis tumors can be enhanced in chickens coinfected with serotype 2 Marek's disease virus, a common vaccine virus. This enhancement requires a genetically susceptible chicken and early infection with avian leukosis virus in addition to serotype 2 Marek's disease vaccination. Because most commercial chicken strains are resistant, and avian leukosis virus infection has been largely eradicated from susceptible stocks, enhancement is not currently recognized as a field problem.
A subclinical disease syndrome characterized by depressed egg production in the absence of tumor formation is more important economically than are deaths from lymphoid leukosis. Chickens with subclinical disease usually shed virus or viral antigen into the albumen of eggs. The pathogenic mechanisms are poorly understood.
Epidemiology of Avian Leukosis in Poultry
Chickens are the natural hosts for all viruses of the leukosis/sarcoma group. These viruses have not been isolated from other avian species except pheasants, partridges, and quail.
Avian leukosis virus is shed by the hen into the albumen or yolk, or both; infection probably occurs after the onset of incubation. Congenitally infected chickens fail to produce neutralizing antibodies and usually remain viremic for life.
Horizontal infection after hatching is also important, especially when chicks are exposed immediately after hatching to high doses of virus (eg, in feces of congenitally infected chicks or in contaminated vaccines). Horizontally infected chickens have a transient viremia followed by antibody production. The earlier the infection, the more likely it is to lead to tolerance, persistent viremia, and tumors.
Other factors known to increase the susceptibility of chickens to horizontal infection include the absence of maternal antibodies and the presence of endogenous retroviruses, especially those associated with the late feathering (K) gene.
Tumors are more frequent in congenital than in horizontal infections; however, many more chickens are exposed horizontally than congenitally. Rates of embryo transmission typically are 1%–10%; virtually all chicks in an infected flock are exposed by contact. Congenital and, in some cases, early horizontal infection can induce permanent carrier states characterized by shedding of virus or antigen into the environment and into eggs. Late infection (ie, inoculation at 12–20 weeks) is unlikely to lead to viral shedding.
Four classes of avian leukosis virus infection are recognized in mature chickens. Mature chickens are designated into 1 of the 4 classes based on their infection status.
No viremia, no antibody (V-A-)
No viremia, with antibody (V-A+)
Viremia, with antibody (V+A+)
Viremia, no antibody (V+A-)
Birds in an infection-free flock and genetically resistant birds in a susceptible flock fall into the category V-A-. Genetically susceptible birds in an infected flock fall into one of the other three categories. Most are V-A+, and a minority, usually < 10%, are V+A-. Most V+A- hens transmit the virus to a varying but relatively high proportion of their progeny.
The virus is not highly contagious compared with other viral agents and is readily inactivated by disinfectants. Transmission can be decreased or eliminated by strict sanitation. After the infection is eradicated, standard disease control and sanitation practices can keep chicken flocks free of the disease.
The role of males in transmission of avian leukosis virus is uncertain. Infected cocks apparently do not influence the rate of congenital infection of progeny but act only as virus carriers and sources of contact or venereal infection to other birds.
Clinical Findings of Avian Leukosis in Poultry
Chickens with lymphoid leukosis have few typical clinical signs. These may include inappetence, weakness, diarrhea, dehydration, and emaciation. Infected chickens become listless before death.
Palpation often reveals an enlarged cloacal bursa and sometimes an enlarged liver.
Infected birds may not necessarily develop tumors; however, they may lay fewer eggs.
Lesions
Diffuse or nodular lymphoid tumors are common in the liver, spleen, and cloacal bursa and are found occasionally in the kidneys, gonads, and mesentery.
Involvement of the cloacal bursa has been considered virtually pathognomonic, although bursal lymphomas are also known to be induced by reticuloendotheliosis virus. Sometimes the bursal tumors are small and observed only after careful examination of the mucosal surface of the organ.
Usually, no enlargement of peripheral nerves is apparent, although such lesions have been noted after experimental inoculation of subgroup J virus.
Microscopically, the tumor cells are uniform, large lymphoblasts. Mitotic figures are frequent. (See lymphocytic infiltration, and images.)

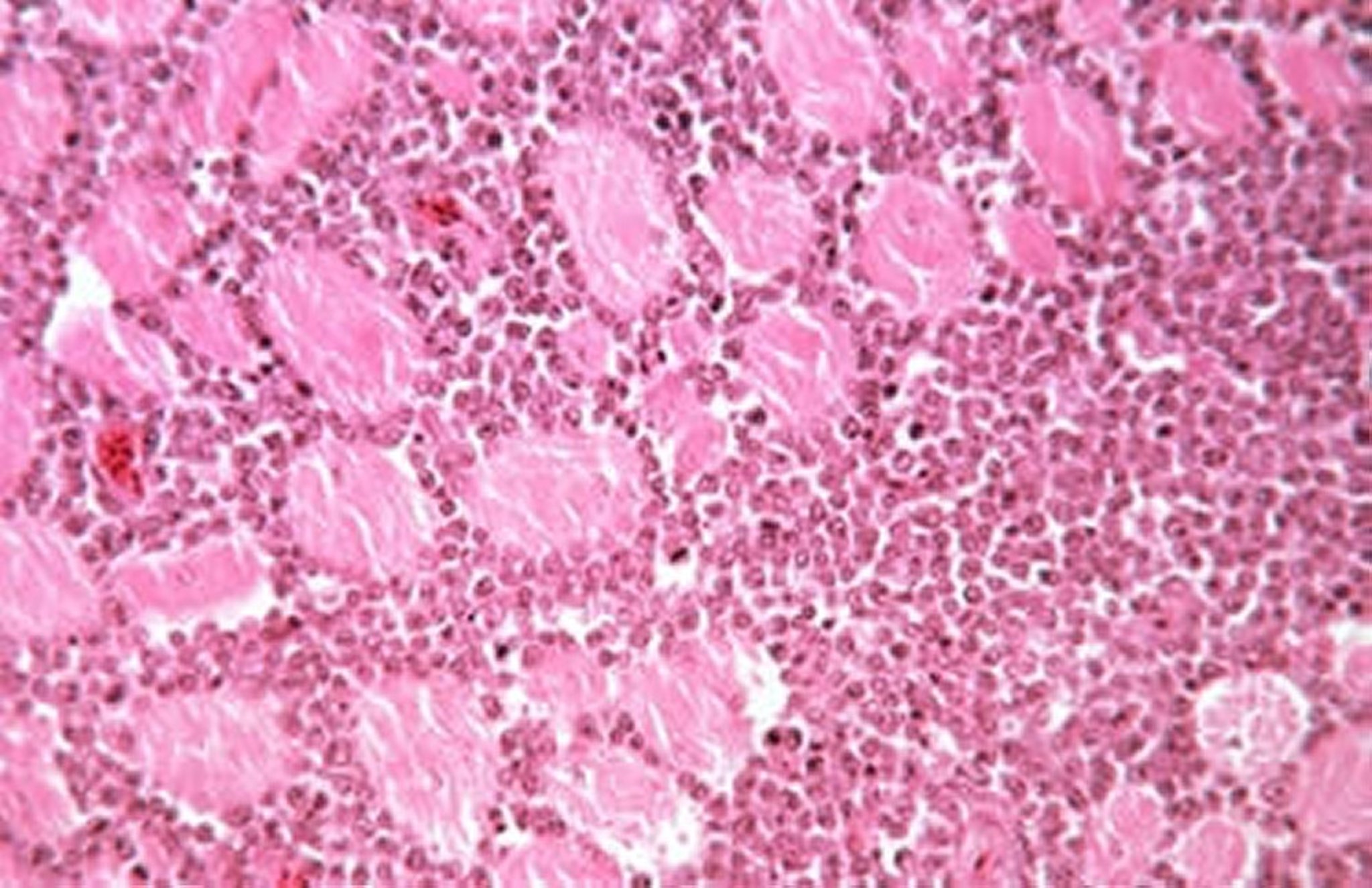
Lymphocytic infiltration in muscle due to lymphoid leukosis. Lymphocytes are uniform in size and morphology, sometimes resembling blast cells. H&E stain; original magnification, 20X.
Courtesy of Dr. Jean Sander.


Lymphocytic infiltration in the liver due to lymphoid leukosis. Lymphocytes are uniform in size and morphology, sometimes resembling blast cells. H&E stain; original magnification, 40X.
Courtesy of Dr. Jean Sander.
Outbreaks of neoplasms other than lymphoid leukosis, such as myelocytomas, hemangiomas, and renal tumors, have also been noted in meat-type chickens infected with subgroup J avian leukosis virus.
Myelocytomas may cause protuberances on the head, thorax, and shanks.
Myelocytomas may occur in the orbit of the eye, causing hemorrhage and blindness.
Hemangiomas may occur in the skin, appearing as “blood blisters,” which may rupture and bleed.
Renal tumors may cause paralysis due to pressure on the sciatic nerve.
Microscopically, in cases of myelocytomas induced by subgroup J avian leukosis virus, the liver shows a massive intravascular and extravascular accumulation of myeloblasts characterized by the presence of cytoplasmic eosinophilic granules.
Most strains of leukosis/sarcoma viruses also induce nonlymphoid tumors (including sarcomas), erythroblastosis, myeloblastosis, myelocytomas, hemangiomas, nephroblastomas, osteopetrosis, and related neoplasms. The nature and frequency of the tumors depend on virus strain, chicken strain, age, dose, and route of infection.
Occasional outbreaks of predominantly one type of tumor are observed in the field. The Rous sarcoma virus, a member of this group, has been widely studied in the laboratory. Each strain usually causes a predominantly neoplastic disease and can be distinguished on the basis of pathogenicity. Some viruses (eg, Rous sarcoma and erythroblastosis viruses) contain a viral oncogene that enables the virus to induce neoplasms within a short incubation period; however, such viruses are rare in the field. Others cannot replicate on their own and require a nondefective helper virus.
Avian leukosis virus infection has also been shown to be associated with the so-called fowl glioma, characterized by cerebellar hypoplasia and myocarditis.
Diagnosis of Avian Leukosis in Poultry
Standard criteria: history and clinical signs, gross pathological findings, and histological examination
Advanced criteria: immunohistochemical testing, standard and quantitative PCR assay, virus isolation, serological testing
Avian leukosis virus is widespread among chickens; therefore, virus detection tests alone, including virus isolation and PCR assay and the demonstration of antigen or antibody, have limited or no value in diagnosing field cases of lymphomas.
Gross characteristics of diagnostic importance include the tumorous involvement of the liver, spleen, or cloacal bursa in the absence of peripheral nerve lesions (see .
The tumors are found in birds > 14 weeks old.
Histologically, the lymphoid cells are uniform in character, large, and contain IgM and B-cell markers on their surface.

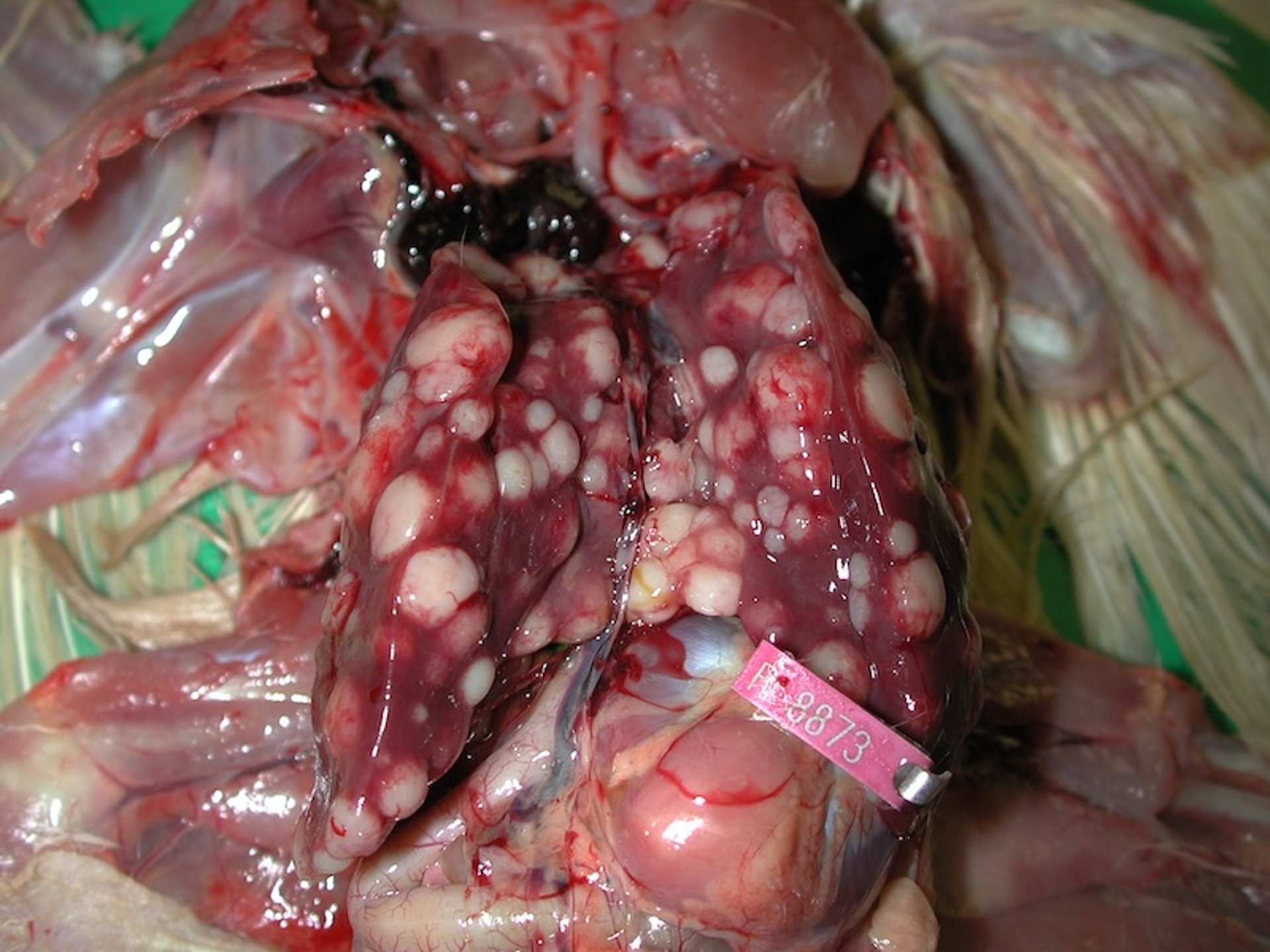
Liver with multifocal tumor nodules in a chicken with lymphoid leukosis.
Courtesy of Dr. John Dunn.
Tumors in avian leukosis can be differentiated from those of Marek's disease on the basis of gross and microscopic pathological changes and by molecular techniques that demonstrate the characteristic clonal integration of proviral DNA into the tumor cell genome with the associated disruption of the c-myc oncogene. Lymphoid leukosis cannot easily be differentiated from B-cell lymphomas due to reticuloendotheliosis virus except by virological assays; however, such tumors probably are extremely rare.
PCR primers specific for detection of each subtype of avian leukosis virus have been developed. PCR assay has been used to detect and characterize avian leukosis virus strains contaminating commercial live virus vaccines of poultry. ELISA kits for detection of antibodies against avian leukosis virus subgroups A, B, and J are available commercially.
Control of Avian Leukosis in Poultry
Eradicating viruses from breeding flocks
Eradication of avian leukosis virus from primary breeding stocks is the most effective means to control avian leukosis virus infection and lymphoid leukosis in chickens. Breeder flocks are evaluated for viral shedding by testing for viral antigens in the albumen of eggs with enzyme immunoassays or by biological assays for infectious virus. Eggs from shedder hens are discarded, so that progeny flocks typically have decreased levels of infection. If raised in small groups, infection-free flocks can be derived with relative ease. These control measures are applied only to primary breeder flocks.
Some breeders favor, and have virtually achieved, total eradication, whereas others favor a decreased level of viral infection. Some chickens have specific genetic resistance to infection with certain subgroups of virus. Although genetic cellular resistance is unlikely to replace the need for decreasing or eradicating the virus, the cellular receptor gene has recently been cloned, and quick molecular assays for viral susceptibility could be developed. Thus far, vaccination for tumor prevention has not been promising.
Key Points
Lymphoid leukosis is a B-cell lymphoma, occurring in chickens ~16 weeks or older.
The cloacal bursa (bursa of Fabricius) is usually involved.
A presumptive diagnosis can be made based on history, clinical signs, gross pathological findings, and histological examination.
For More Information
Nair V, Gimeno I, Dunn J, et al. Leukosis/Sarcoma Group. In: Swayne DE, ed. Boulianne M, Logue CM, McDougald LR, Nair V, Suarez DL, associate eds. Diseases of Poultry. 14th ed. Wiley; 2019:587-625.
Witter RL, Gimeno IM, Pandiri AR, Fadly AM. Tumor Diagnosis Manual: The Differential Diagnosis of Lymphoid and Myeloid Tumors in the Chicken. American Association of Avian Pathologists; 2010.
References
Zhao Z, Rao M, Liao M, Cao W. Phylogenetic analysis and pathogenicity assessment of the emerging recombinant subgroup K of avian leukosis virus in South China. Viruses. 2018;10(4):194. doi:10.3390/v10040194





