

Meningitis, encephalitis, and encephalomyelitis are terms used to describe inflammatory conditions of the meninges, brain, or brain and spinal cord, respectively. These inflammatory processes frequently occur concurrently, with the terms meningoencephalitis and meningoencephalomyelitis used. Although such conditions have long been associated with bacteria, viruses, fungi, rickettsial agents, and parasites, more recent findings in both people and animals are starting to also implicate chemical agents and immune-mediated processes, with genetic predispositions proposed for some of these latter conditions. Depending on the causal agent, the extent and speed of onset, and location of the inflammation, clinical signs can vary from subtle to dramatic, and therapies can have varied successes. Accurate diagnosis is dependent on quality neurologic (including ocular) examinations and supported by CSF analysis and imaging, in particular MRI. In unsuccessful cases, the importance of postmortem examinations cannot be underestimated, because historically, many zoonotic agents have manifested as meningoencephalitis in people and animals.
Inflammation of the meninges (meningitis) and inflammation of the brain (encephalitis) are seen in animals and often manifest concurrently (meningoencephalitis). Many of the inflammatory diseases of the CNS of animals are diffuse, involving both the brain and spinal cord (encephalomyelitis and meningoencephalomyelitis). Because many inflammatory processes are disseminated throughout the CNS at the time of clinical observation, differentiation between meningeal-only inflammation vs extension of disease into the neuropil is often difficult to make antemortem. Thus, from a clinical standpoint, any one of these conditions may be the case in an animal with an inflammatory condition of the CNS.
In animals with meningoencephalitis or meningoencephalomyelitis, the clinical signs of meningitis often precede those of encephalitis and may remain the predominant feature of the illness. This is especially apparent in meningitis involving neonates.
Causes of meningitis, encephalitis, and meningoencephalitis include bacteria, viruses, fungi, protozoa, rickettsia, parasite migrations, chemical agents, and idiopathic or immune-mediated diseases.
In ruminants, generally bacterial infections are more common than other causes of meningitis or encephalitis. In species other than ruminants, especially adult animals, viruses, protozoa, rickettsia, and fungi are as or more frequent causes of meningitis or encephalitis than are bacteria.
The appearance of many etiologic agents such as arboviruses, certain rickettsia, and bacteria) are seasonal. Age is an important consideration for bacterial meningitis associated with neonatal sepsis. Even within species (eg, production animals), there can be a difference in risk factors. For instance, sporadic bovine encephalomyelitis , caused by Chlamydia pecorum, and thromboembolic meningoencephalitis, caused by Histophilus somni, usually occur in feeder beef cattle (6 mo to 2 yr old) and not replacement dairy calves unless managed on feedlots.
Vaccine history is also an essential factor in consideration of differential diagnoses in animals with clinical signs of inflammation of the CNS, especially those caused by viruses.
Etiology and Pathogenesis of Meningitis, Encephalitis, and Encephalomyelitis in Animals
The incidence of inflammation in the brain and meninges is relatively low compared with that of inflammation in other organs; however, the etiologies are quite variable, with bacterial infections more common in farm animals while viral and parasitic agents are reported more in dogs, cats, and horses. Although for many years the brain and, to some extent, the meninges were considered sanctuaries against infectious agents, it is now known that many mechanisms exist for agents to invade the meninges, brain, and spinal cord. These mechanisms vary from direct adjacent invasion such as from sinus or ear infections, to bloodborne infections that compromise vascular endothelial cells, the trafficking of viruses within blood cells through an intact or compromised blood-brain barrier, or trafficking of viruses and certain bacteria along nerves. It is also worth noting that invasion of the meninges allows access to CSF, a dynamic, continually flowing medium that facilitates bacterial and viral replication and travel both within the central canal and ventricle system and, via the subarachnoid spaces, to levels of the lumbar cord and cauda equina.
Bacterial Infections

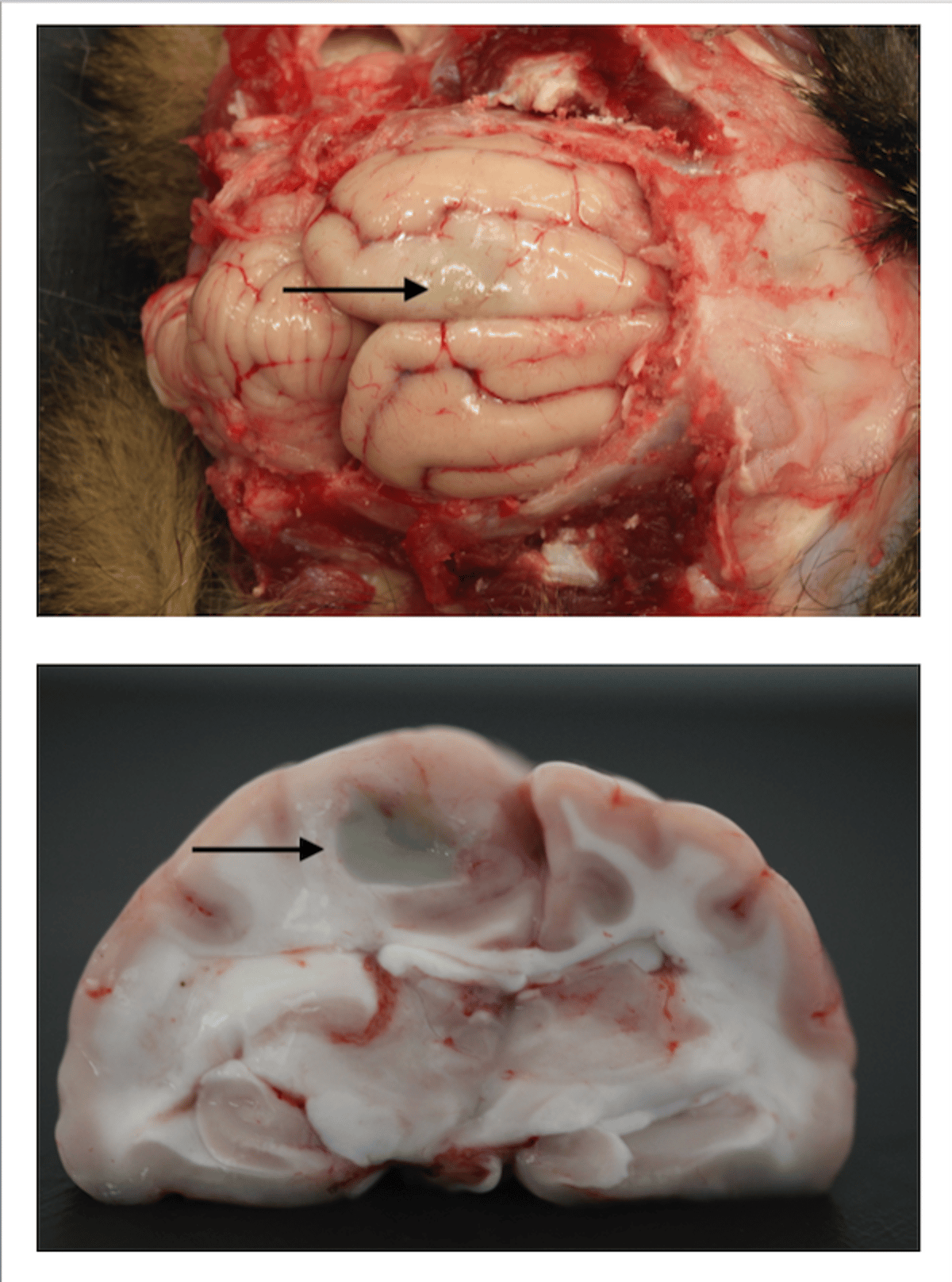
Surface (top) and cross-sectional (bottom) view of cerebral hemispheres of a cat revealing a suppurative process induced by a puncture wound to the skull, speculated to be due to a cat bite.
Courtesy of Brian Cloak.
Bacteria can arrive in the CNS either as a result of local trauma or by direct extension, as occurs with puncture wounds from fighting (cats), tail biting (pigs), sinusitis after dehorning (cattle), otitis media or interna(Mycoplasma bovis in calves), spinal column inflammation (salmonellosis in cattle) or migrating grass awns in hunting dogs. Hematogenous spread, as septic emboli from suppurative lesions in other locations of the body (endocarditis or arthritis in cattle or dogs, navel-ill in ruminants) or as bacteremia that leads to CNS vascular damage (Histophilus somni in feedlot cattle), is also well documented as a mode of bacterial entry to the CNS. Whereas these latter lesions lead to blood-brain barrier damage, penetration of an intact blood-brain barrier can also occur by cells harboring intracellular bacteria such as Mycobacterium spp, leading to granulomatous inflammatory lesions. Penetration of CNS by axonal transport, a prominent feature of viral infections, is also noted in listeriosis.
Bacterial infections can result in the development of brain abscesses, granulomas, foci of infarction, or hemorrhage and focal to widespread, moderate to severe meningitis. Brain abscesses and granulomas occur in a number of disorders but are particularly associated with peripheral chronic inflammatory conditions that deliver septic emboli to the brain (bacterial endocarditis in cattle and dogs), deep puncture wounds from bites or migrating foreign bodies, or in association with intracellular pathogens such as Mycobacterium spp (in many species) or Rhodococcus equi infection in foals. Although abscesses are typically relatively random in location (eg,adjacent to sites of direct trauma,at the junction of gray and white matter in the brain for septic emboli), pituitary abscesses are more common in ruminants and thought to originate from bacterial invasion of the rete mirabile (intracranial carotid mirabile) surrounding the pituitary gland.

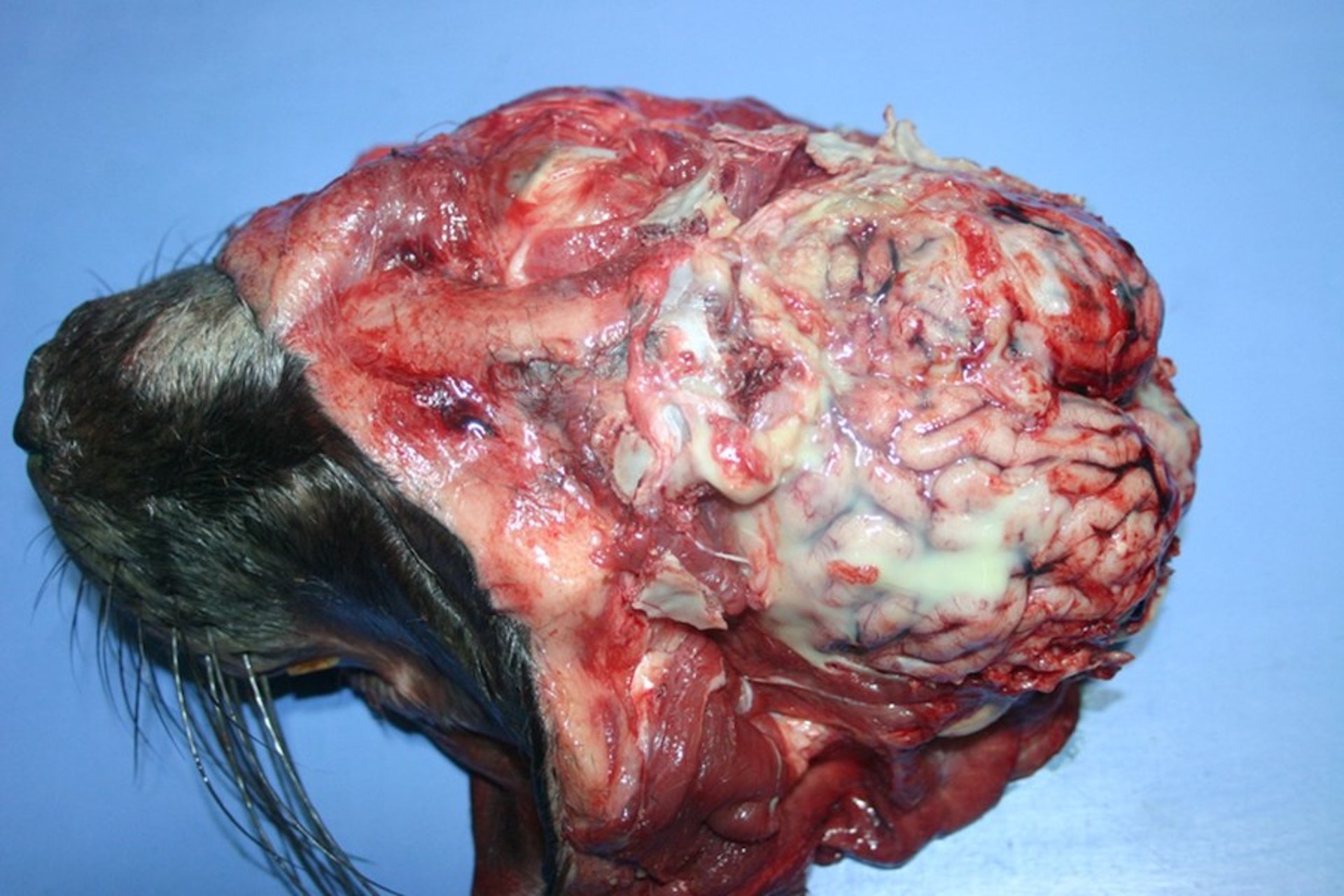
Bacterial meningitis in an Atlantic Grey Seal (Halichoerus grypus). The presence of significant pus is typical of a bacterial etiology.
Courtesy of Brian Cloak.
Bacterial meningitis or meningoencephalitis develops more commonly in food animals than other species, is likely to be sporadic, and is most common in neonatal animals as a sequela of gram-negative septicemia associated with navel-ill. Thus, many CNS manifestations are accompanied by lesions in other parts of the body. Of note are the navel-ill infections: streptococcal infections of ruminants and pigs, E coli, particularly of ruminants, and Actinobacillus equulis in foals. Across species, various aerobic bacteria (Pasteurella multocida, Mannheimia haemolytica,Staphylococcus spp, Actinomyces spp, and Nocardia spp) and anaerobic bacteria (Bacteroides spp, Peptostreptococcus anaerobius, Fusobacterium spp, Eubacterium spp, and Propionibacterium spp) have also been isolated from lesions of bacterial meningitis and encephalitis.
Sporadic meningoencephalitis in horses, donkeys, and mules has been associated with Mannheimia haemolytica and Actinomyces, Klebsiella, and Streptococcus spp. In Lyme (Borrelia burgdorferi) disease–endemic areas, where the agent is known to induce joint, cardiac, and CNS (meningoencephalitis) inflammation in people, neurologic infection in dogs and horses is likely to be reflective of a meningitis and/or encephalitis. Other nonneonatal, hematogenously derived infections that are considered to manifest as epidemics of meningoencephalitis with high mortality include thromboembolic meningoencephalitis (Histophilus somni) of feedlot cattle, Chlamydophila pecorum in yearling cattle, Glässer disease of weaner pigs (Haemophilus parasuis), and Haemophilus agni septicemia in feeder lambs.
Encephalitis as a primary lesion with a predilection for the brain stem and accompanied by a meningitis is the most common manifestation of listeriosis, which is caused by Listeria monocytogenes. Infection is seen in cattle, sheep, and goats and less commonly in horses. Incidentally, several species of Mycoplasma cause encephalitis in goats (M mycoides), poultry (M gallisepticum), cats (M felis), dogs (M edwardii) and rodents (M pulmonis).
It is noteworthy that some bacterial infections may be considered zoonotic, such asCapnocytophaga canimorsus, a commensal, gram-negative bacterium from the mouth of dogs and cats. It is known to induce meningitis in elderly and immune-compromised patients, and Pasteurella multocida can induce meningitis in infants. Although there is limited information on the pathogenesis of Elizabethkingia (formerly Chryseobacterium) meningoseptica, this bacterium causes meningitis in newborn and immunocompromised people and animals.
Viral Infections
Many viruses cause meningitis, encephalitis, and meningoencephalitis. Viral infections typically cause nonsuppurative inflammation, devoid of neutrophils, pus, or significant necrosis, other than that caused by death of individual infected cells. Often, the extent of inflammation does not correlate well with the intensity of clinical signs. Several viruses are specifically neurotropic or exhibit predilection for the CNS, causing a fulminant or fatal encephalitis, the most notorious being rabies virus infection. Although rabies viruses primarily spread to the CNS transaxonally, several other common DNA (adenoviruses, herpesviruses, parvoviruses) and RNA (bunyavirus, lentiviruses, morbilliviruses, alphaviruses, flaviviruses) viruses are likely to enter the brain via the blood-brain barrier (within cells) and exhibit high neuropathogenesis once within the CNS. A third mechanism is viruses that induce CNS vascular damage, either by inducing a vasculitis in their viremic phase, resulting in loss of vascular integrity and subsequent hemorrhages and edema or inducing a chronic granulomatous inflammation centered on and around blood vessels, the latter observed in feline infectious peritonitis (dry form) caused by feline coronavirus. Malignant catarrhal fever, caused by ovine herpesvirus 2, and canine herpesvirus encephalitis, caused by canine herpesvirus 1, represent classic examples of viral vasculitis in cattle and dogs, respectively.
Although many viral causes of encephalitis and meningitis are known, the list continues to expand, and situations continue to be seen in which clinical signs and lesions suggest a viral etiology yet an agent has not been identified. For instance, in people, only 30%–50% of suspected cases of viral encephalitis are confirmed by the detection of the etiologic agent in CSF samples via PCR. In addition, a causative agent has yet to be identified in several encephalitic conditions in birds, mink, cattle, fish, and people in which the histologic features, together with clinical, epidemiologic, and pathologic observations, support a viral etiology. Add to this the fact that because, historically, a number of zoonotic conditions manifest as encephalitis or meningoencephalitis (rabies, West Nile virus, Nipah virus, Hendra virus, and controversially Borna disease virus), there is a constant need to be vigilant and accurately determine the cause of group outbreaks of encephalitis.
In veterinary medicine, louping ill virus is a common encephalitis of sheep and cattle, with occasional reports in horses, deer, grouse, dogs, and people. Louping ill, a flavivirus transmitted by ticks, likely uses hematogenous and transaxonal routes to enter the brain and infect neurons, eliciting neural necrosis and a diffuse inflammatory response with an accompanying meningitis. Rabies (Rhabdovirus family) and Aujeszky disease (pseudorabies caused by Herpesviridae family), transmitted by biting or by eating infected meat, respectively, are highly infectious to a multitude of species and use nerve axons to enter the CNS and exert their effects by also infecting neurons and causing variable levels of inflammation.

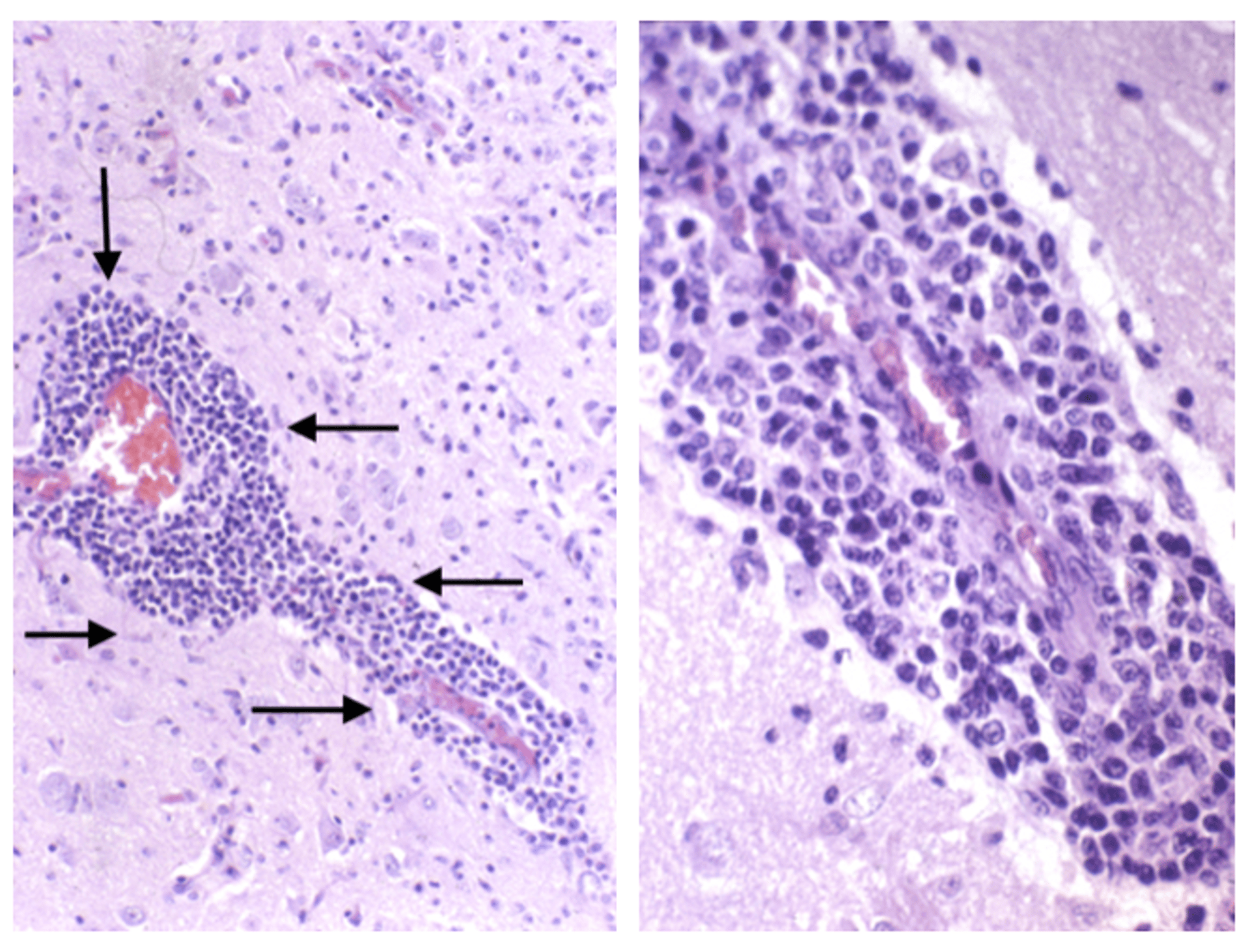
A perivascular cuffing of inflammatory cells (mainly lymphocytes and plasma cells) in the brain, a hallmark of a viral or immune-mediated inflammatory process. Right: closeup.
Courtesy of Brian Cloak.
Viruses should be considered the prime causes of group outbreaks of meningitis, encephalitis, and meningoencephalitis, particularly in cattle, sheep, pigs, and horses. Full necropsies of recently dead animals can considerably help establish diagnoses. Because many histopathologic hallmarks of viral infections exist, evaluations of CNS tissues are crucial to the identification of the causal agent, which can be further confirmed by molecular analysis of tissues, which has in many instances replaced the need for more labor-intensive tissue culture processes. Meningoencephalitis may only be one manifestation of the viral infection, which may also involve lesions in other organs (lentiviral infections). The presence of ticks (flaviviruses) or mosquitoes (West Nile virus, Japanese encephalitis virus in people and pigs, and Eastern, Western, and Venezuelan encephalitis in horses) may significantly influence the direction of the investigation and the potential zoonotic implications of the outbreak.
Infections Caused by Parasites

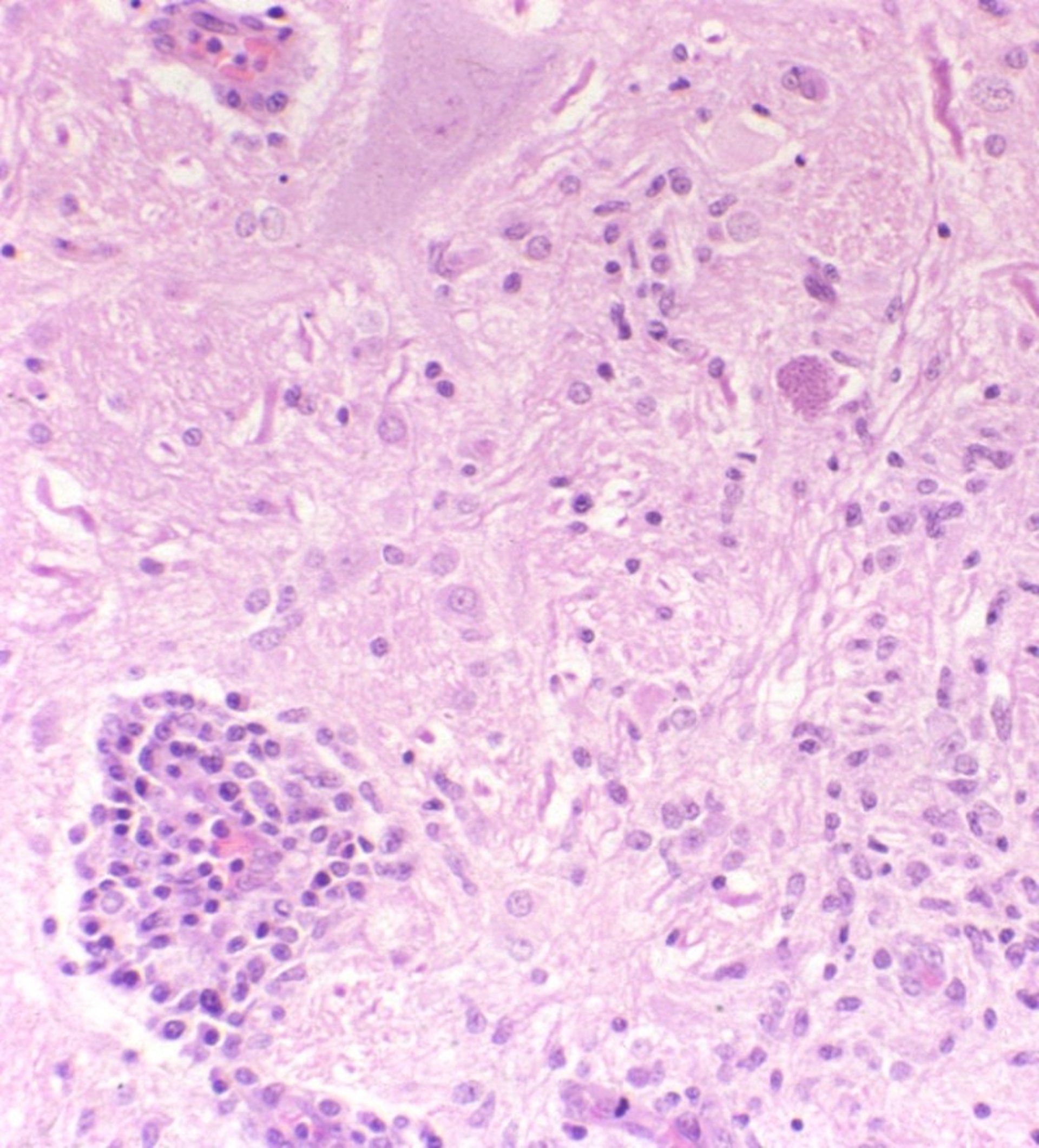
Brain inflammation associated with protozoal cysts (Neospora caninum) in a dog.
Courtesy of Brian Cloak.
Many parasitic agents can cause meningoencephalitis in large and small animals. Neuropathogenic protozoa include Toxoplasma gondii, Neospora caninum, and Encephalitozoon cuniculi in dogs and cats. E cuniculi can cause encephalomyelitis in rabbits. Sarcocystis neurona and N hughesii are important causes in horses, with Trypanosoma spp important in equids outside of the USA. A wide variety of protozoa can infect and cause severe CNS disease in adult cattle, including Babesia bovis, Theileria parva (theileriosis), and Trypanosoma spp, whereas N caninum and T gondii can cause congenital encephalitis in calves. Free-living amoebae, Naegleria fowleria, Acanthamoeba spp, and Balamuthia mandrillaris are associated with amoebic meningoencephalitis in dogs.
Aseptic suppurative or eosinophilic meningoencephalitis associated with aberrant migration of parasites throughout the CNS can develop in a number of animal hosts. In dogs and cats, CNS infections have been reported with Dirofilaria immitis, Toxocara canis, Ancylostoma caninum, Cuterebra spp larva, and Taenia spp. A wide variety of nematodes have been reported to cause severe meningoencephalitis in horses, including Setaria spp, Habronema spp, Strongylus spp, Halicephalobus gingivalis, and Angiostrongylus cantonensis (the rat lungworm). In cattle, migrating Setaria and Hypoderma larva are commonly implicated. Recently, A cantonensis has been identified as a cause of eosinophilic meningoencephalitis in nonhuman primates. Parelaphostrongylus tenuis is especially important in goats and llamas.
Fungal Infections
Pathogenic fungi, including Coccidioides immitis, Blastomyces dermatitidis, and Histoplasma capsulatum, can cause meningoencephalitis. Opportunistic invasion with Cryptococcus neoformans and Aspergillus spp has also been described in several mammalian species. Rarely, other fungi, such as Candida spp, Cladosporium trichoides, Paecilomyces variotii, Geotrichum candidum, and dematiaceous fungi (Bipolaris sp and Alternaria spp) cause meningoencephalitis. Unicellular plants, Prototheca wickerhamii and P zopfii, can also produce an eosinophilic meningoencephalomyelitis in dogs, cattle, and horses.
Idiopathic/Immune Mediated
Several idiopathic meningoencephalitides are recognized in dogs, likely driven by immune processes that may have genetic, infectious, or a combination of both triggers at their core. Granulomatous meningoencephalomyelitis is a relatively common but sporadic CNS disease that most often affects young to middle-aged dogs. Although small-breed females are more typically affected, there is not a clear familial or genetic basis as a driver for this immune-mediated encephalitis.


Greyhound meningoencephalitis, a breed-specific neuroinflammatory condition. This greyhound is displaying ventroflexion of the neck.
Courtesy of Brian Cloak.
In contrast, a number of breed-specific neuroinflammatory conditions exist, including those observed in Pug, Maltese, Yorkshire Terrier, Pointer, and Greyhound dogs. All conditions display variable levels of inflammation, which appear viral-like in the young adult Greyhounds and are accompanied by significant necrosis in young adult Pug, Maltese, and Yorkshire Terrier dogs. In mature Pointer dogs, the lesion is a pyogranulomatous meningoencephalomyelitis.
A steroid-responsive meningitis, with variable levels of neutrophils, affecting mainly young (less than 2-year-old), large-breed dogs, and a severe necrotizing vasculitis and meningitis syndrome have been documented in Beagles, Bernese Mountain Dogs, German Short-haired Pointers, and Nova Scotia Duck-tolling Retrievers. Both conditions have been identified as possible immunologic disorders with a hereditary predisposition. An eosinophilic meningoencephalitis that has been described in adult dogs (Golden Retrievers, Rottweilers, and South African Boerboels) and rarely in cats is also believed to have an immunologic basis.
Clinical Findings of Meningitis, Encephalitis, and Encephalomyelitis in Animals
In the early stages of inflammatory diseases of the CNS, nonlocalizing clinical signs are frequent. In dogs, for example, meningitis can easily be mistaken for intervertebral disc extrusion, polyarthritis, pleuritis, pancreatitis, or pyelonephritis. In horses, initial clinical signs can appear as lameness, myositis, vertebral instability, or even colic. In foals, extreme hyperexcitability and irritability can be an early indication of sepsis in the CNS. Cattle can demonstrate anorexia, depression, and bizarre behavior, including strange vocalization due to peripheral nerve damage.
The usual signs of meningitis are fever, hyperesthesia, neck rigidity, and painful paraspinal muscle spasms. Dogs and occasionally horses display this syndrome acutely and occasionally chronically, without clinical signs of brain or spinal cord involvement. However, in diffuse meningoencephalitis due to any agent, depression, blindness, progressive paresis, cerebellar or vestibular ataxia, opisthotonos, cranial nerve deficits, seizures, dementia, agitation, and depressed consciousness (including coma) can develop, depending on the rapidity of onset, pathology, and location of the lesions. Visual deficits, neck pain, seizures, behavioral disturbances, ataxia, weakness, cranial nerve deficits, and depression may be seen in either focal or disseminated CNS disease.
In neonatal infections, omphalophlebitis, polyarthritis, and ophthalmitis with hypopyon can accompany the CNS inflammation. Because of its unusual pathogenesis and development of asymmetric brain lesions, listeriosis often causes asymmetric vestibular dysfunction, with head tilt and circling, in addition to other cranial nerve deficits such as facial and pharyngeal paralysis. In histophilosis of cattle, the nervous signs tend to be peracute, with sudden collapse and profound depression of consciousness (stupor or coma); fever and limb stiffness may be the only signs detectable in the prodromal stages. In sporadic bovine encephalomyelitis, calves demonstrate incoordination that can progress to recumbency and opisthotonos.
Lesions
Gross lesions are extremely variable depending on cause and location and whether the disease is diffuse or multifocal. Pathologic changes characteristic of meningitis include diffuse infiltration of leukocytes into the leptomeninges. This can be mild in flaviviral infections, or visible to the eye as an increased opacity with thickened meninges (in suppurative bacterial meningitis), or be a series of multifocal hemorrhages if the organisms damage vascular endothelium.
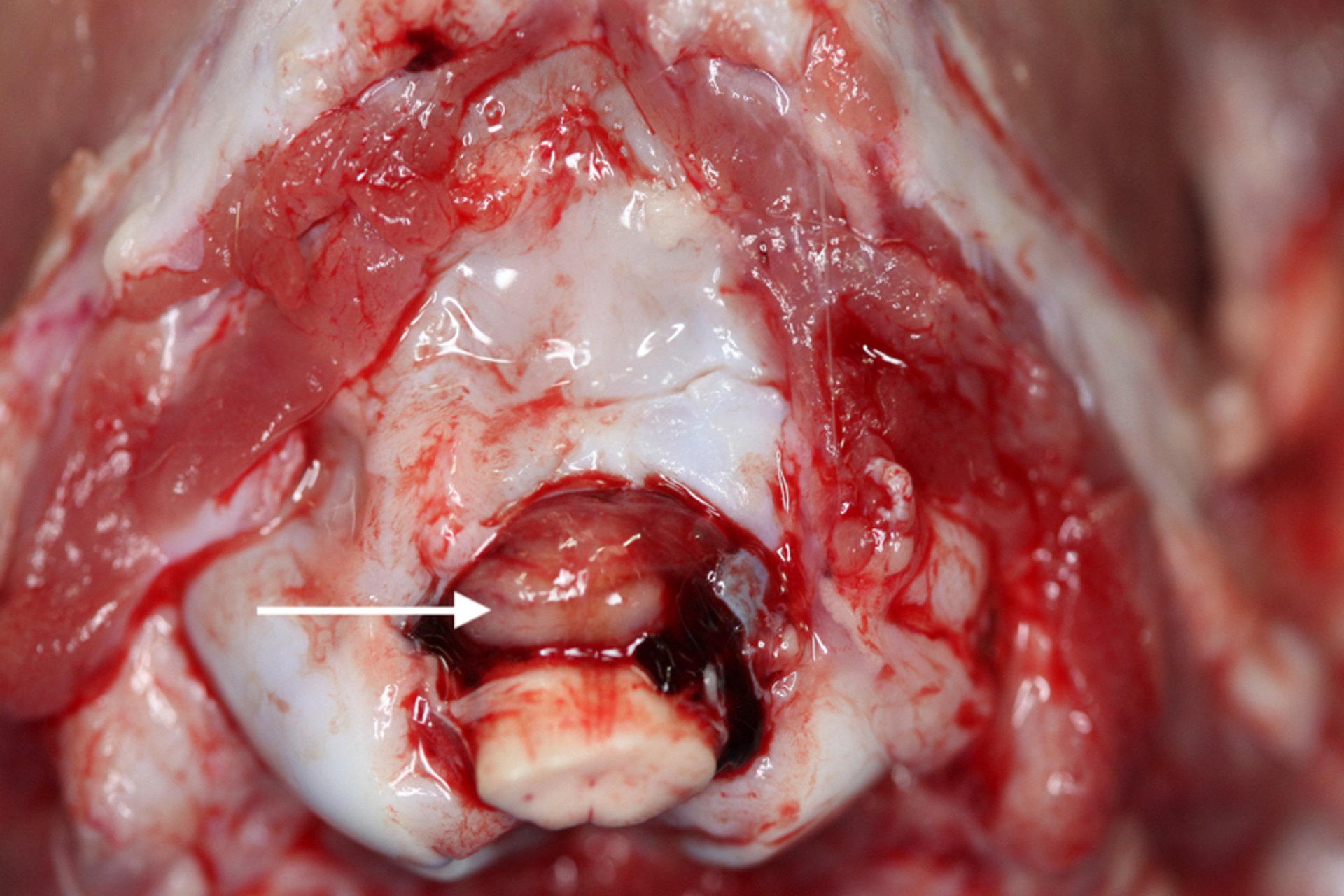
Herniation of the caudal part of the cerebellum (coning of the cerebellum) at the foramen magnum, suggestive of intracranial pressure due to brain edema or space-occupying lesions.
Courtesy of Brian Cloak.
Depending on the insult and the agent, varying levels of necrosis may be present in addition to inflammation. This may take the form of individual cell death, microabscesses, or extensive necrosis with cavitations or large abscesses. There may also be regions of demyelination, hemorrhage, and edema. Grossly, this may translate to observing swollen and/or hemorrhagic brain tissue and/or herniation of brain tissue. Regions of cavitation or abscessation can often been seen grossly. The identification of causal agents cannot be confirmed from gross examination, however, apart from focal cryptococcal lesions and the cysts formed by the cestode Coenurus cerebralis, the larval stage of Taenia multiceps.
Histopathologic examination allows for the identification of the hallmarks that separate viral, bacterial, parasitic, and fungal infections, often allowing for the visualization of agents, or in the case of viruses, some of their "footprints" such as inclusion bodies, the nature of the cell infiltration, and the levels and location of necrosis.
Diagnosis of Meningitis, Encephalitis, and Encephalomyelitis in Animals
History and clinical evaluation, supported by CSF evaluation and imaging (typically MRI)
In large outbreaks, necropsies coupled with PCR, immunohistochemistry, and serology are necessary
CBCs and serum biochemical profiles often are unremarkable in CNS infections. The analysis of CSF is the most reliable and accurate means to identify an encephalitis, meningitis, or meningoencephalitis. CSF should be collected whenever history, species, or breed predisposition suggests meningitis or encephalitis, or whenever clinical signs indicate a disseminated or multifocal CNS disorder. Without CSF analysis, an animal exhibiting back or neck pain with an increase in rectal temperature may be misdiagnosed.
Adult large animals and dogs with bacterial meningitis and encephalitis or with steroid-responsive suppurative meningitis typically have a marked neutrophilic pleocytosis in the CSF. The protein content of the CSF is usually also significantly increased (>100 mg/dL), with an increase in the globulin component of CSF.
Rickettsial infections most often cause a mild to moderate mononuclear pleocytosis, although Rocky Mountain spotted fever can cause neutrophilic inflammation secondary to vasculitis.
In foals with suspected bacterial meningitis, the CSF has an increased protein content, and even slight increases in WBCs in the CSF are significant (>10 WBC/mcL). Any neutrophils observed on cytology in CSF from a foal warrant treatment with antimicrobials that can obtain high therapeutic levels in the CNS.
Viral infections and listeriosis typically produce a mild to moderate mononuclear (usually lymphocytic) pleocytosis in CSF, with an associated increase in protein levels. However, the CSF is normal in rabies virus infections. Herpesviral infections cause markedly increased proteins and xanthochromia (yellow to reddish discoloration) without dramatic increase in cell count. Feline infectious peritonitis in cats and Eastern equine encephalitis in horses are exceptions and can cause markedly high neutrophil counts. In feline infectious peritonitis, a markedly high protein concentration (>200 mg/dL) can also be seen.
Parasitic and fungal meningoencephalitides cause eosinophilic or occasionally a highly degenerate neutrophilic pleocytosis. Granulomatous inflammations usually induce moderate to high cell numbers and increased protein in the CSF. The cell population is predominantly mononuclear or a mixed population of neutrophils and mononuclear cells. Distinguishing a granulomatous infection due to a fungal or protozoal organism from granulomatous meningoencephalitis is often difficult. The necrotizing encephalitides typically cause a mild increase in CSF mononuclear cells and protein concentration.
Occasionally, bacteria are seen on cytologic examination of the CSF and identified by Gram stain. Successful culture of bacteria from CSF is more likely in large animals than in dogs. In some cases, serial blood cultures are more successful, especially in foals. Fungi and occasionally protozoa have been identified in CSF, but serology is usually necessary to confirm mycotic and protozoal infections in vivo. Many of these diseases are fatal, and final identification is made at postmortem with in situ identification of the organism.
For premortem etiologic identification, agent-specific testing is recommended; however, most agents, once in the CNS, are not detectable by direct testing through culture or nucleic acid–based testing of body fluids. Serologic testing is available for most viral encephalitides and, in particular, for arboviruses the most reliable test examines IgM in a single sample.
Paired serum is required for IgG-based tests, especially those confounded by vaccination. Although CSF analysis is rewarding in terms of clinical pathology, detection of a pathogen within the CSF can also be unreliable depending on the location and pathogen load within the CNS. Culture of the CSF will often yield growth of the organism in bacterial meningitis; however, the detection rate is often < 40% for many viruses. Detection of antibody within the CNS can be nonspecific if there is leakage through the blood-brain barrier. IgM detection is likely a more reliable indication of intrathecal antibody production. Most confirmatory testing is performed postmortem if the animal dies.
Treatment of Meningitis, Encephalitis, and Encephalomyelitis in Animals
Antimicrobials or other specific therapy and supportive care
Depending on the clinical signs of the animal, supportive therapy is likely required in addition to specific therapies focused on the etiologic agent or the mechanism driving the CNS inflammation. Other than for animals with the probable immune-mediated, steroid-responsive inflammatory CNS diseases and for animals with meningoencephalitis caused by rickettsia, protozoa, and certain bacteria, the prognosis is guarded. Although anti-inflammatory therapy can reduce the immune drive observed in the immune-mediated and breed-associated meningoencephalitic disorders, in many instances fatalities ultimately occur. The case fatality rate in calves with bacterial meningitis has been reported to be up to 100%; however, the case fatality rate in foals is much lower.
Appropriate use of antibiotics, according to culture or serology results, is basic to successful therapy. Relapses are common, and prolonged therapy is often necessary. Correction of failure of passive transfer is critical in neonatal large animals. Broad-spectrum antibacterials that can penetrate the blood-brain barrier should be selected, and bactericidal drugs are preferred over bacteriostatic agents. Recommended drugs include ampicillin, metronidazole, tetracyclines, potentiated sulfonamides, fluoroquinolones, and third-generation cephalosporins; higher than normal dosages may be necessary to achieve and maintain adequate concentrations in the CNS. In farm animals, selection of drugs must be based not only on drug efficacy but also on whether the available drug is appropriate for use in a food animal.
For viral infections, the case-fatality rate varies. The most lethal viral infections are rabies (100%) in all mammals, Eastern equine encephalomyelitis in horses (85%–100%), and distemper virus in dogs (50%). Availability of antivirals is limited, and cost can be prohibitive. The most commonly treated viral infection of the CNS is likely that of the neurotropic form of equine herpesvirus 1; however, the prognosis is guarded in recumbent horses.
Mycotic infections of the CNS have been treated successfully in people, but results in veterinary medicine are less rewarding. Treatment with itraconazole or fluconazole may be of benefit, but longterm therapy is required and relapses are frequent. Protozoal infections (eg, toxoplasmosis, neosporosis, sarcocystosis) may respond to a potentiated sulfonamide (trimethoprim, pyrimethamine and sulfonamides). These are commonly used in combination with clindamycin in small animals. However, relapse may occur because of the inability to clear encysted organisms from the CNS. Antiprotozoal medications have been approved for use in horses, such as the triazine analogues, including diclazuril and ponazuril. In balantidiasis, a disease in working donkeys, secnidozole has been shown to decrease fecal cyst counts, which should theoretically decrease the risk of development of CNS disease.
Glucocorticoids are usually contraindicated in animals with meningitis or meningoencephalitis with an infectious etiology; however, a high-dose, short-term course of dexamethasone or methylprednisolone may control life-threatening complications such as acute cerebral edema and impending brain herniation. Immunosuppressive doses of corticosteroids are required for successful therapy of the idiopathic CNS inflammations seen in dogs.
Radiation therapy and immunomodulatory drugs have been used to treat granulomatous meningoencephalitis.
Supportive care should be specific for the needs of the individual animal and may include analgesics, anticonvulsants, fluids, nutritional supplementation, and physical therapy.
Key Points
Neurologic examinations and CSF analysis are important starting points in the evaluation of meningoencephalitic conditions in animals.
In group outbreaks, postmortem examinations, including histopathology and molecular analysis, must be considered to help identify the etiologic agent.
Potential zoonotic implications must be considered, particularly in group outbreaks, in certain locations, and in situations for which the etiologic agent cannot be determined.
For More Information


