



West Nile virus (family Flaviridae, genus Orthoflavivirus) is a widespread, mosquito-borne pathogen maintained in nature in bird-mosquito transmission cycles. Dead-end hosts include horses, humans, and numerous other mammalian species, with fatal disease a potential outcome of infection in some species. Reptiles, including alligators, are also susceptible to disease. It is primarily a pathogen of wild and exotic birds, in which it can cause fatal neurological disease. Acute infections can be diagnosed by suggestive neurological clinical signs (eg, ataxia, weakness, hind limb paresis, head tilt, tremors, torticollis, seizures) or histological examination combined with virus isolation, RT-PCR assay, or immunohistochemical testing. Some patients may recover with supportive care, such as fluid administration and assisted feeding; however, prognosis generally is poor in cases of severe neurological disease.
West Nile virus (WNV), reported on all continents except Antarctica, is the most widespread of the flaviviruses. It was one of the first arthropod-borne viruses (ie, arboviruses) identified to infect humans; it was first isolated from a febrile Ugandan woman in 1937.
Outbreaks initially were reported in Israel in the 1950s, followed by France and Russia in the 1960s and Belarus and Ukraine in the 1970s. The role of mosquitoes in virus transmission and of wild birds as virus reservoir (amplifying) hosts were described in the 1950s.
WNV was first associated with disease in domestic avian species in 1997, when flocks of young geese in Israel were affected with a neurological disease. In August 1999, the disease appeared for the first time in the Western Hemisphere when wild and zoo birds, horses, and humans died in the northeastern US, initially in the New York City area.
Domesticated fowl are generally much less susceptible to WNV-associated disease than other avian species, although some geese and ducks may develop fatal disease.
Susceptibility to WNV-associated disease varies greatly among wild bird species, ranging from subclinical infection to death (with mortality rates approaching 100%). Species within the family Corvidae (eg, crows and jays) are considered the most susceptible to severe morbidity and death due to WNV, although many raptors (eg, owls, hawks, and eagles), some songbirds, waterfowl, and other birds may develop fatal disease.
Among wild game birds, the greater sage-grouse and ruffed grouse develop disease and die most frequently, whereas other game birds generally are less susceptible to severe disease. Also see section on equine arboviral encephalomyelitis.
Etiology of West Nile Virus in Birds
West Nile virus is in the family Flaviviridae, which contains the genera Orthoflavivirus, Hepacivirus, Pestivirus, and Pegivirus:
Orthoflaviviruses include WNV, Japanese encephalitis virus (JEV), dengue virus (DENV), and yellow fever virus (YFV).
Hepaciviruses include hepatitis B and C viruses.
Pestiviruses primarily affect hoofed mammals.
Pegiviruses have not been clearly associated with disease but cause persistent infections in a wide range of mammals.
The Orthoflavivirus genus contains more than 70 viruses, which are further classified into tickborne and mosquito-borne virus groups. Some mosquito-borne viruses within this genus (eg, WNV and JEV) comprise the Japanese encephalitis serocomplex (ie, the encephalitic clade), while others (eg, DENV, YFV) are in the hemorrhagic fever clade (ie, the nonencephalitic clade).
Epidemiology of West Nile Virus in Birds
Geographical distribution of mosquito-borne flaviviruses such as West Nile virus largely depends on the preferred habitats of competent mosquito vectors, with mainly Culex spp mosquitoes transmitting encephalitic flaviviruses in the Northern Hemisphere. WNV is endemic in many countries of Africa; Asia; southern Europe; and North, Central, and South America. Epidemics in human populations may occur at infrequent and unpredictable intervals, often based on environmental conditions that alter mosquito activity.
Most flaviviruses are transmitted by mosquito or tick vectors, although some have no currently recognized vectors. WNV has been detected in at least 43 mosquito species in the US, among which C pipiens generally is considered most important. The virus also has been isolated from hard (ixodid) and soft (argasid) tick species in regions of Europe, Africa, and Asia where WNV is endemic, although the likelihood and thus importance of tickborne WNV transmission is unknown.
WNV is maintained in natural transmission cycles between mosquito vectors and avian reservoir hosts. Numerous wild bird species (eg, crows, robins, sparrows, grackles, hawks, owls, and others) reach sufficient viremia titers to infect mosquitoes (based on laboratory studies in C pipiens mosquitoes), while viremia titers in chickens and domestic turkeys rarely reach this level.
Because WNV is primarily transmitted by mosquito, disease detection and outbreaks are often seasonal (ie, from midsummer to mid-to-late fall) in temperate regions, while they may occur year-round in tropical regions.
Transmission of West Nile Virus in Birds
The principal route of West Nile virus transmission is by the bite of an infected mosquito (primarily Culex spp). The predominant mosquito vector species vary geographically, and preferred host species differ among mosquito species.
Bird-to-bird transmission also has been demonstrated experimentally when birds are housed in close quarters, likely via ingestion of shared virus-contaminated food and water sources because the virus can be shed orally and cloacally.
Direct ingestion of infectious WNV (fluid, mosquitoes, or infected tissues) also resulted in experimental infection in some birds. Raptor species (ie, birds of prey) may be infected via ingestion of infected prey. Although ingestion is likely an uncommon and not epidemiologically meaningful transmission route, it remains a possibility in domestic birds (eg, poultry) in high-density housing with shared food and water sources.
Clinical Findings of West Nile Virus in Birds
In general, birds that are susceptible to West Nile virus–associated disease can exhibit a variety of nonspecific clinical signs (eg, emaciation, dehydration, abnormal molt or feather loss, weakness, recumbency, drooped head, anorexia, lethargy, fluffed feathers) and neurological clinical signs (eg, ataxia, head tilt, nystagmus, tremors, hind limb paresis, seizures, blindness). See images of WNV infections in American crow, great horned owl, long-eared owl, and red-tailed hawk.


Courtesy of Dr. Nicole Nemeth.


Courtesy of Dr. Nicole Nemeth.


Courtesy of Dr. Sigrid Ueblacker.


Courtesy of Dr. Sigrid Ueblacker.
Young (immature) birds of many species generally are more susceptible to disease, exhibiting higher viremia titers than do more mature (ie, adult) birds. Very young (eg, 1 day post-hatch) chickens experimentally infected with WNV exhibited fluffed feathers, weakness, and lethargy before death. Older chicks and adult chickens and turkeys are not known to develop WNV-associated disease and appear highly resistant to disease. Further, domestic chickens often seroconvert without ever having shown clinical signs of disease, and experimental infections have failed to induce disease.
Among other domesticated or semidomesticated fowl, some breeds of geese and ducks are clinically affected by WNV infection. Similarly, some species of wild waterfowl and other aquatic birds are susceptible to West Nile disease. The most susceptible wild birds belong to the corvid family (eg, crows, jays, and ravens), although numerous other passerine (eg, sparrows, grackles, and finches), raptor (eg, owls, hawks, eagles, and falcons), and psittacine (eg, parrots, parakeets, conures) species as well as a few Galliformes (eg, ruffed grouse, greater sage-grouse) also are susceptible to fatal disease.
Lesions
There are no pathognomonic gross lesions; however, irregular pale streaks, irregular foci or mottling on the epicardium and myocardium, or a uniformly pale heart may occur, as may splenomegaly and hepatomegaly (see images of WNV with pale heart and splenomegaly). Blood vessels within the meninges may be congested; however, this is a nonspecific and inconsistent finding. Emaciation may also be observed; however, in many cases, disease is acute and birds are in fair to good nutritional condition.

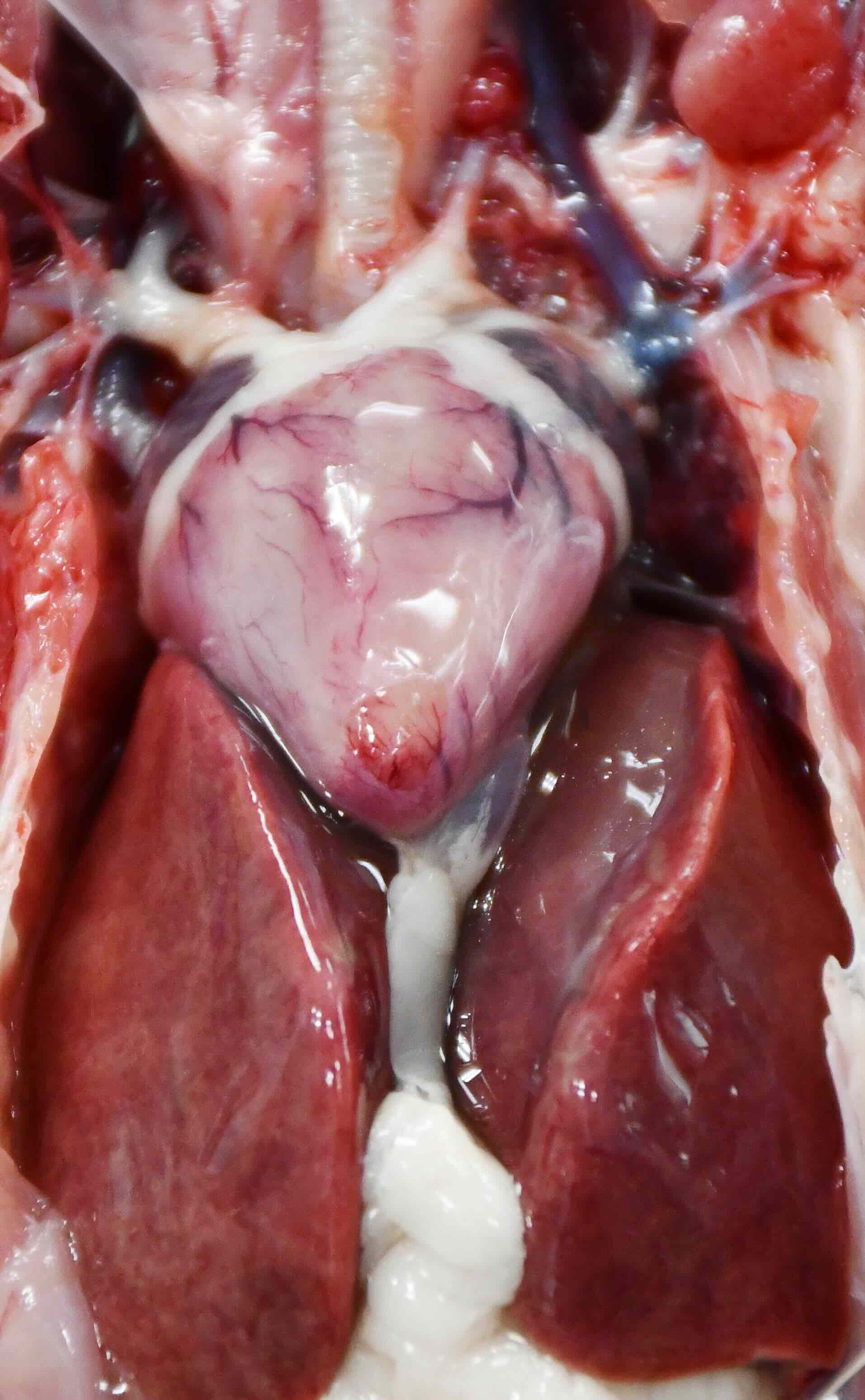
Courtesy of the Southeastern Cooperative Wildlife Disease Study.


Courtesy of the Southeastern Cooperative Wildlife Disease Study.
Histopathological findings may include nonsuppurative (lymphoplasmacytic) myocarditis, encephalitis, endophthalmitis, nephritis, hepatitis, adrenalitis, perineuritis (eg, in the myenteric plexus), perivasculitis, and arteritis.
In addition, myocardial, adrenal gland, and pancreatic necrosis may occur either without (ie, peracute to acute) or concurrent to inflammation. In acute cases, the myocardium may have subtle to extensive degeneration and necrosis of myofibers, while more chronic cases may be associated with lymphoplasmacytic myocarditis. Similarly, brain lesions may be minimal early in infection; however, as infection progresses, brain lesions may develop to lymphoplasmacytic (often perivascular) meningoencephalitis, gliosis, and neuronal degeneration.
Diagnosis of West Nile Virus in Birds
Antemortem: serological testing, virus isolation, or RT-PCR assay of blood or swabs
Postmortem: virus isolation, RT-PCR assay, or immunohistochemical labeling in conjunction with histological examination of tissues
West Nile virus–associated disease in birds can clinically manifest as nonspecific (eg, emaciation, dehydration, abnormal molt or feather loss, anorexia, lethargy, fluffed feathers) or neurological disease (eg, ataxia, head tilt, nystagmus, tremors, hind limb paresis, seizures, and blindness). In some cases, death may occur without premonitory clinical signs.
Diagnosis generally is by virus isolation or RT-PCR assay of blood or other tissues, which can be pooled to maximize detection. RT-PCR assay using swabs from the oropharyngeal cavity (less so the cloacal cavity) may also prove useful in acute cases. In birds that die of acute infection, high viral titers generally occur in tissues, blood, and oropharyngeal and cloacal secretions. Postmortem diagnosis can also be confirmed by histological examination in conjunction with immunohistochemical labeling.
Serological testing (ELISA) is not widely commercially available; however, demonstration of 4-fold or higher increase in anti-WNV antibody titers (acute and convalescent; ie, collected 2–4 weeks apart) also indicates recent infection. A single seropositive sample merely demonstrates previous infection because anti-WNV antibodies likely persist lifelong in many avian species; thus, timing of infection and causation of disease cannot be determined with a single seropositive result.
The plaque reduction neutralization test is the gold standard for WNV serological testing; however, it is labor-intensive because the closely related St. Louis encephalitis virus (SLEV) must be ruled out by paired serum titrations due to cross-reactivity.
Preferred postmortem tissues for diagnosis of recent WNV infection by virus isolation and RT-PCR assay are heart, spleen, kidney, and brain; pooling small pieces of each tissue for testing may increase the chances of detection. Brain is useful but may provide false-negative results if tested as the sole tissue, especially in acute infections.
Tissues must be homogenized before virus isolation or RT-PCR assay. RT-PCR assay is the most commonly used diagnostic test for WNV; virus isolation is less commonly performed on a commercial basis because it requires biosafety containment facilities. Tissues may have infectious virus for up to ~1–2 weeks after initial infection (and blood for ~1 week), after which the virus often is rapidly cleared from tissues in surviving birds.
Immunohistochemical labeling also can be used to confirm infection via viral antigen detection in formalin-fixed paraffin-embedded tissues. This may involve a variety of tissues; however, heart, kidney, adrenal gland, spleen, and brain can be most useful, depending on the timing after infection. Evaluation and testing of tissues with acute lesions is preferred. In addition, immunohistochemical labeling of mononuclear cells circulating in the blood may be present and is indicative of infection.
Differential diagnoses for West Nile disease in birds include other encephalitic viruses (eg, highly pathogenic avian influenza viruses and eastern equine encephalitis virus); parasitic infections such as Sarcocystis, Toxoplasma, and Baylisascaris spp; bacterial infections by agents such as Riemerella anatipestifer, Streptococcus gallolyticus, and Erysipelothrix, Listeria, and Salmonella spp; viscerotropic velogenic Newcastle disease; avian paramyxovirus-1; and avian bornavirus.
Trauma, some toxins (eg, ionophores, organophosphates and carbamates, lead), and CNS and multicentric neoplasia (eg, lymphoma) also may cause nonspecific or neurological clinical signs in birds and are additional differential diagnoses that must be considered in conjunction with husbandry and environmental conditions.
Treatment, Prevention, and Control of West Nile Virus in Birds
Supportive care
Mosquito control
Vaccination for limited protection
Treatment of WNV-associated disease is primarily supportive and includes parenteral fluids, assisted feeding, supplemental heat if indicated, and confinement in a padded cage (in case of seizures). Such measures are typically limited to high-value birds such as those in zoological or other captive settings and endangered species.
Mosquito control is an essential component of programs to prevent West Nile virus or any other mosquito-borne disease. Unfortunately, this is difficult to implement in large backyard and rural environments because of the distances that mosquitoes can fly or be carried by prevailing winds.
Standing water and similar insect-breeding sites in the vicinity of densely populated backyard and other outdoor flock settings, as well as avian farms, should be treated with larvicides or emptied when possible, especially during seasons of known mosquito breeding. Poultry houses and other captive bird enclosures (including zoos and some rehabilitation settings) should be constructed to be mosquito-proof (eg, via use of fine-mesh screens or solid construction that prevents mosquito passage through walls, windows, or doors).
In some areas, local or regional mosquito control programs perform aerial insecticide spraying, which targets both adult and larval mosquitoes. Because WNV and many other arthropod-borne viruses are zoonotic, public health agencies may be involved in mosquito control and management in some areas. Spraying of some chemicals near where birds are housed may be detrimental to their respiratory health, and thus, nontoxic alternatives should be considered in such cases.
Although there are no commercially available WNV vaccinations licensed for use in birds, numerous formulations (eg, killed, canarypox-vectored, plasmid-mediated DNA) have been tested and used in a variety of avian species. Use is generally limited to highly valued captive individuals or flocks (eg, threatened or endangered species recovery programs, zoological parks, educational collections).
Immune protection resulting from these vaccinations is variable and should not be assumed; booster immunization often is necessary. Further, passive transfer of maternal antibodies has been documented in some avian species and may interfere with vaccination of immature birds.
Zoonotic Risk of West Nile Virus From Birds
West Nile virus is zoonotic, with the primary risk of infection being via mosquito bite. WNV is an important cause of neurological disease in humans, with ~1/150 (0.67%) WNV-infected humans developing severe neurological disease, including encephalitis. Similar to recommendations for birds and other animals, the best prevention strategy for humans is avoidance of mosquito bites.
Key Points
In general, poultry and domestic turkeys are resistant or have low susceptibility to WNV-associated clinical disease, in contrast to some species of wild Galliformes (eg, some grouse) and other avian taxa.
Susceptibility to severe disease and death among wild bird species varies; however, corvids (crows, jays) often die most readily and frequently, while many raptors (eg, owls, hawks, eagles) and some psittacines (eg, parrots, parakeets, conures) may also develop fatal disease.
Due to lack of specific treatment options, prevention of WNV infection by avoiding mosquito contact with birds is key.
Vaccination may be used in some birds, but response is variable, and no commercially available vaccines are licensed for use in birds.
For More Information
West Nile virus. Centers for Disease Control and Prevention. Updated June 13, 2023. Accessed February 2, 2023.
West Nile virus. USDA National Invasive Species Information Center. Accessed February 2, 2023.
Where in the United States has West Nile virus been detected in wildlife? United States Geological Survey. Accessed November 25, 2023.
Guy JS. Arbovirus infections. In: Swayne DE, ed. Boulianne M, Logue CM, McDougald LR, Nair V, Suarez DL, associate eds. Diseases of Poultry. 14th ed. Wiley; 2019:507-515.
Kunkel MR, Mead DG, Ruder MG, Nemeth NM. Our current understanding of West Nile virus in upland game birds. Wildl Soc Bull. 2022;46:e1269. doi:10.1002/wsb.1269
Nemeth N. West Nile virus in raptors. In: Fowler ME, Miller RE, eds. Fowler’s Zoo and Wild Animal Medicine: Current Therapy. Vol. 7. Elsevier Saunders; 2012:329-335.
Nemeth NM, Oesterle PT. West Nile virus from an avian conservation perspective. Int Zoo Yearb. 2014;48(1):101-115. doi:10.1111/izy.12031
Palmieri C, Franca M, Uzal F, et al. Pathology and immunohistochemical findings of West Nile virus infection in Psittaciformes. Vet Pathol. 2011;48(5):975-84. doi:10.1177/0300985810391112






