

Coccidiosis is a protozoal disease causing diarrhea, ,weight loss and decreased production in poultry. It can be fatal. Prevention is key and is achieved with use of anticoccidials or vaccination. Diagnosis is by fecal flotation to detect oocysts, often in combination with characteristic necropsy findings.
Coccidiosis is caused by protozoa of the phylum Apicomplexa, family Eimeriidae. Most species affecting poultry belong to the genus Eimeria and infect various intestinal sites. The disease course is rapid (4–7 days) and is characterized by parasite replication in host cells with extensive damage to intestinal mucosa. Coccidia in poultry are generally host-specific, and the different species infect specific portions of the intestine. However, in game birds, including quail, the coccidia may infect the entire intestinal tract. In poultry, game birds reared in captivity, and wild birds, coccidiosis occurs worldwide. (Also see Cryptosporidiosis.)
Etiology
Coccidia are almost universally present in poultry-raising operations, but clinical disease occurs only after ingestion of relatively large numbers of sporulated oocysts by susceptible birds (eg, those that are immunosuppressed and/or with concurrent disease). Both clinically infected and recovered birds shed oocysts in feces, which contaminate feed, dust, water, litter, and soil. Oocysts may be transmitted via equipment and personnel (eg, shoes) as well as the presence of insects (eg, flies) and rodents. Fresh oocysts are not infective until they sporulate; under optimal conditions (70°–90°F [21°–32°C] with adequate moisture and oxygen), this requires 1–2 days. The prepatent period is 4–7 days. Sporulated oocysts may survive for long periods, depending on environmental factors. Oocysts are resistant to some disinfectants commonly used around livestock but are killed by freezing or high environmental temperatures. (Also see Coccidiosis.)
Pathogenicity of coccidiosis is influenced by host genetics, nutritional factors, concurrent diseases, age of the host, and species of the coccidium. Eimeria necatrix and E tenella are the most pathogenic in chickens, because schizogony occurs in the lamina propria and crypts of Lieberkühn of the small intestine and ceca, respectively, and causes extensive hemorrhage. E kofoidi and E legionensis are the most pathogenic in chukars, and E lettyae is most pathogenic in bobwhite quail. Several Eimeria species are pathogenic in pheasants, particularly E phasiani and E colchici. Most species develop in epithelial cells lining the villi.
Protective immunity usually develops in response to moderate and continuing infection. True age-related immunity does not occur, but older birds are usually more resistant than young birds because of earlier exposure to infection.
Clinical Findings and Lesions
Signs of coccidiosis range from decreased growth rate to many sick birds, withsevere diarrheaand high mortality. Decreased feed and water consumption, weight loss, and decreased egg production, may accompany outbreaks. Mild infections which would otherwise be classed as subclinical, may potentially lead to secondary infection, particularly Clostridium spp infection. Birds that survivors severe infections typically recover in 10–14 days but may never recover full growth and production. The lesions are almost entirely in the intestinal tract and often have a distinctive location and appearance that is useful in diagnosis.
Chickens

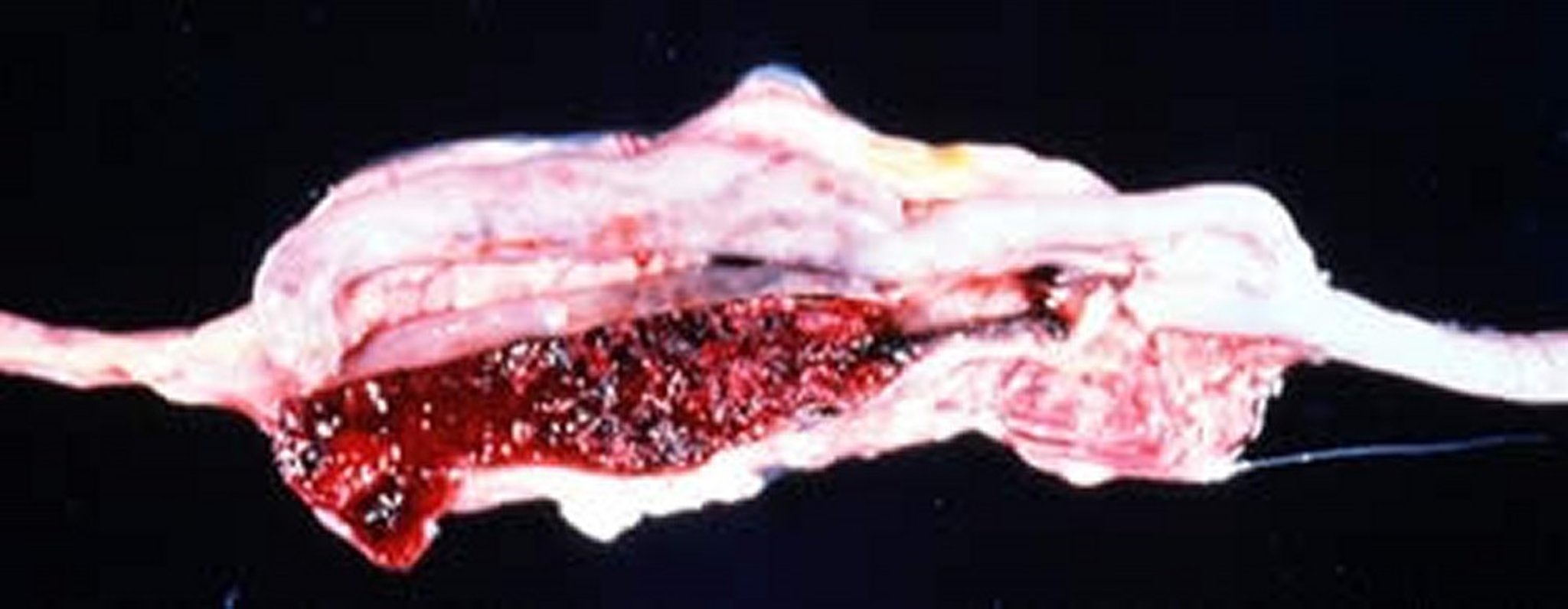
Gross lesions due to E tenella infection, with frank hemorrhaging into cecal pouches in a broiler chicken.
Courtesy of Dr. Jean Sander.

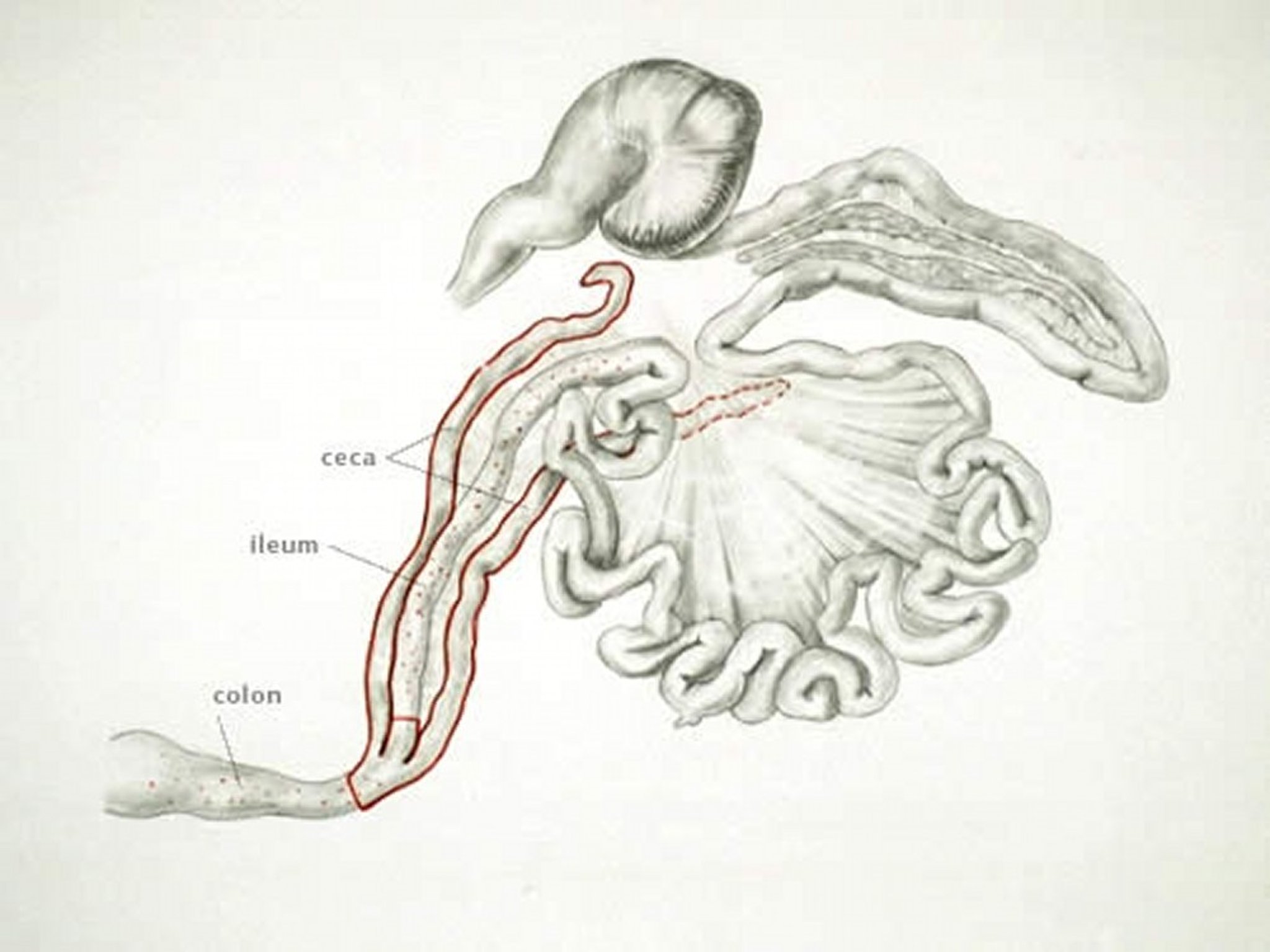
Anatomic drawing illustrating the site of infection with E tenella in poultry
Anatomic drawing by Dr. Gheorghe Constantinescu.
E tenella infections are found only in the ceca and can be recognized by accumulation of blood in the ceca. Cecal cores, which are accumulations of clotted blood, tissue debris, and oocysts, may be found at necropsy in birds surviving the acute stage.

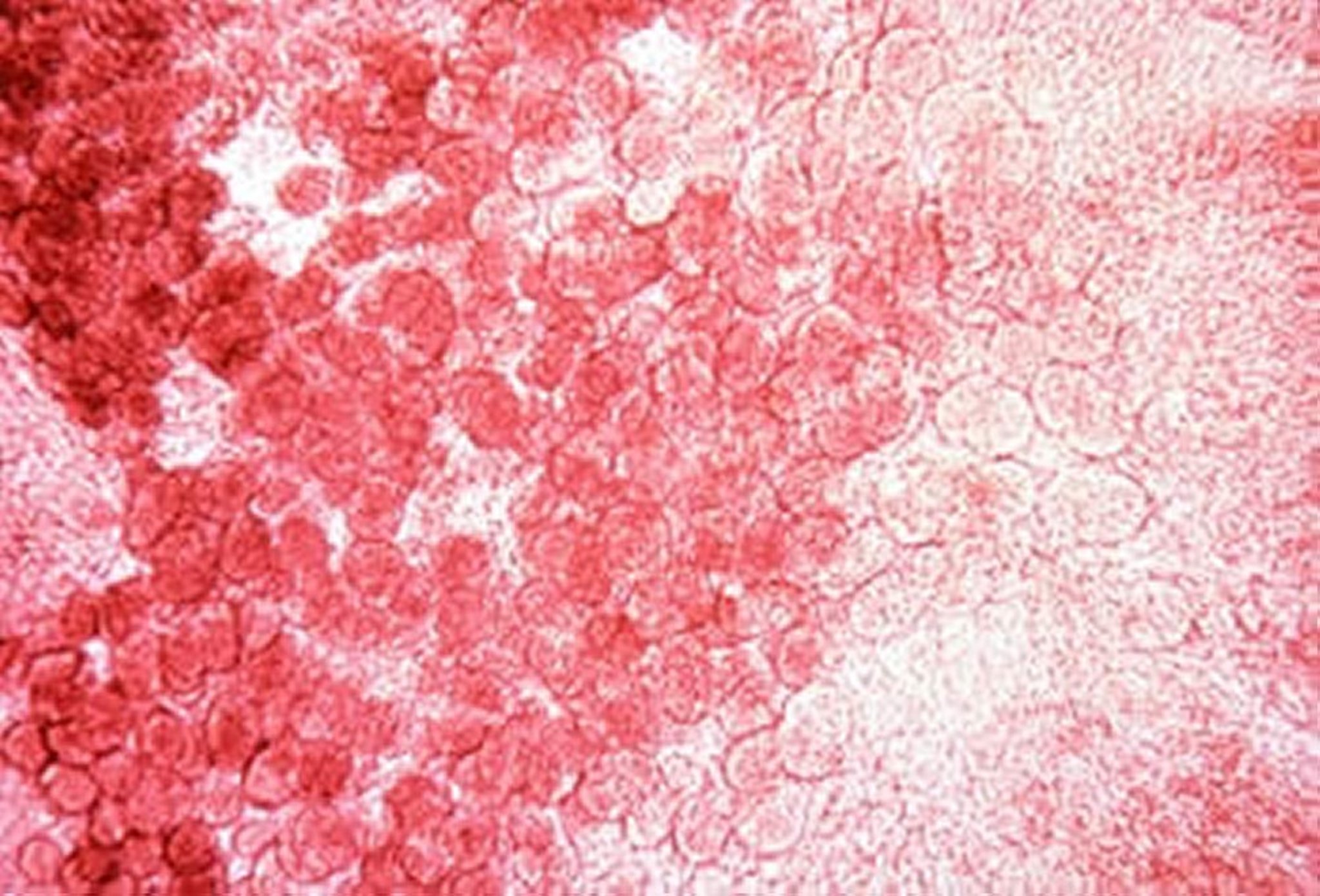
Photomicrograph of a scraping of the midgut showing developmental stages of E necatrix. Feulgen stain, 40X.
Courtesy of Dr. Jean Sander.

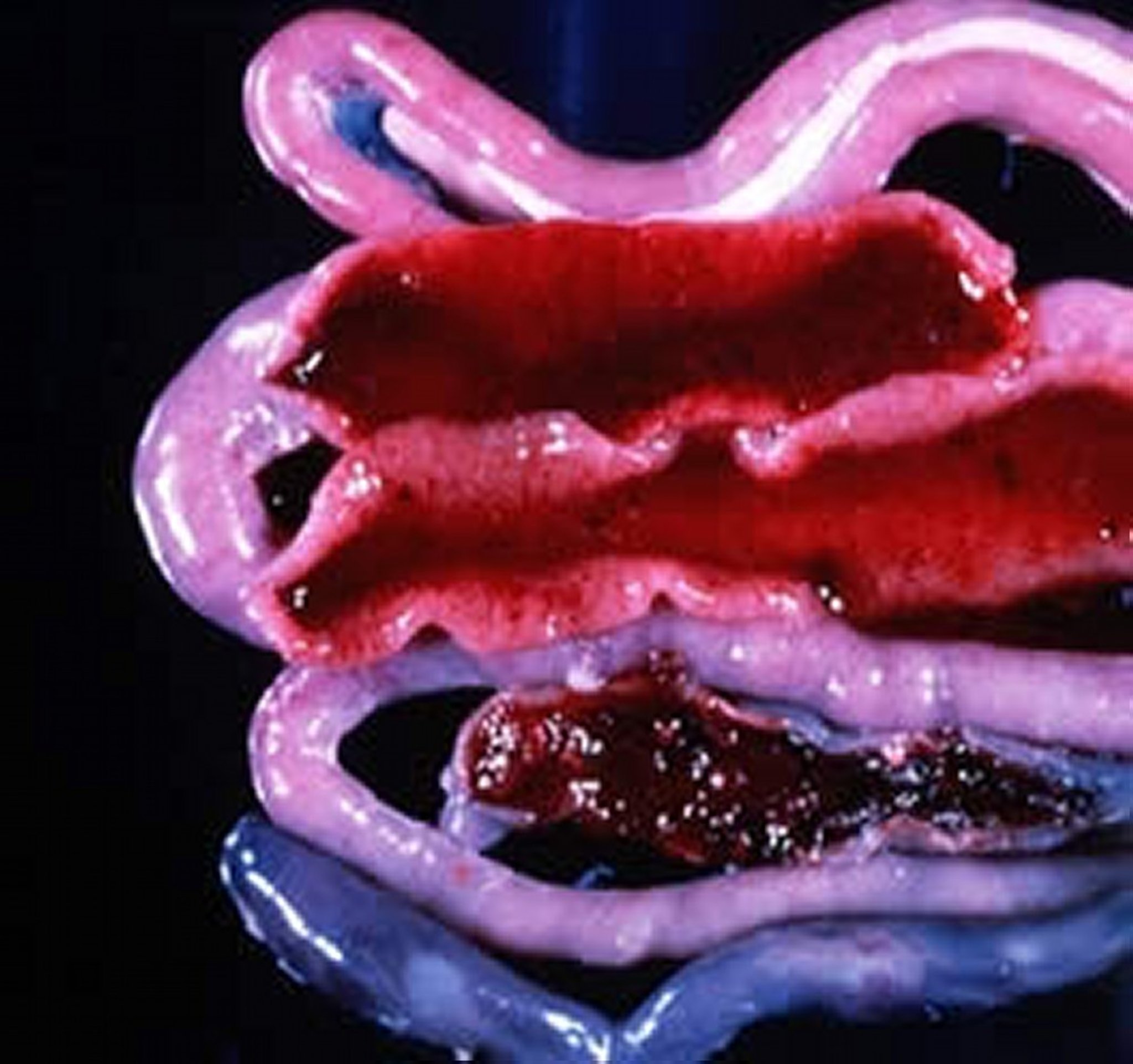
Gross necropsy image showing the midgut of a chicken infected with E necatrix; frank hemorrhaging is present.
Courtesy of Dr. Jean Sander.

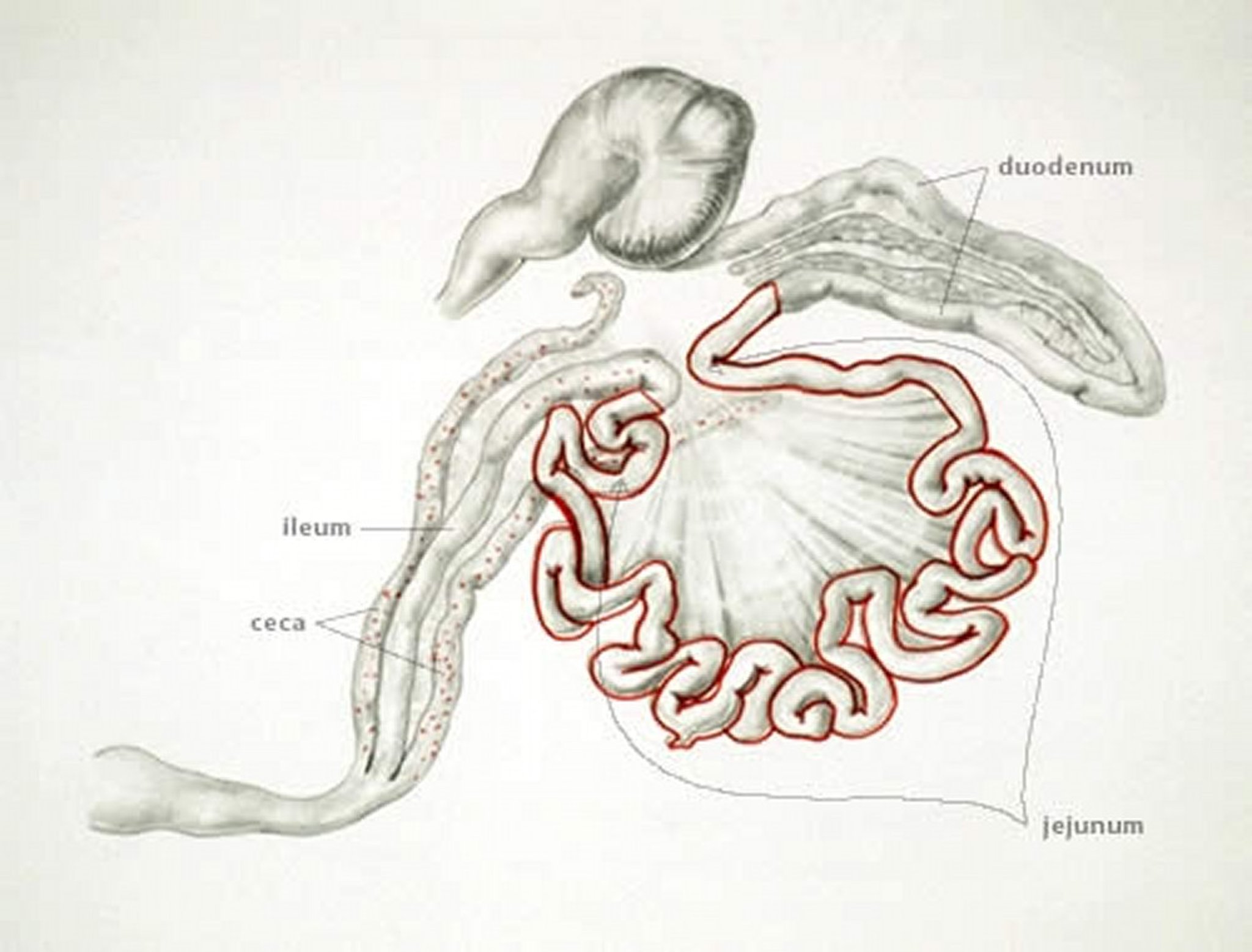
Anatomic drawing illustrating the small intestine as the site of asexual replication and ceca as site of sexual replication of E necatrix in poultry.
Anatomic drawing by Dr. Gheorghe Constantinescu.
E necatrix produces major lesions in the proximal and mid portions of the small intestine. Small, white spots, usually intermingled with rounded, bright- or dull-red spots of various sizes, can be seen on the serosal surface. This appearance is sometimes described as “salt and pepper.” The white spots are diagnostic for E necatrix if clumps of large schizonts can be demonstrated microscopically. In severe cases, the intestinal wall is thickened, and the infected areas are dilated to 2–2.5 times the normal diameter. The lumen may be filled with blood, mucus, and fluid. Fluid loss may result in marked dehydration. Although the damage is in the small intestine, the sexual phase of the life cycle is completed in the ceca. Oocysts of E necatrix are found only in the ceca. Because of concurrent infections, oocysts of other species may be found in the area of major lesions, complicating the diagnostic process.

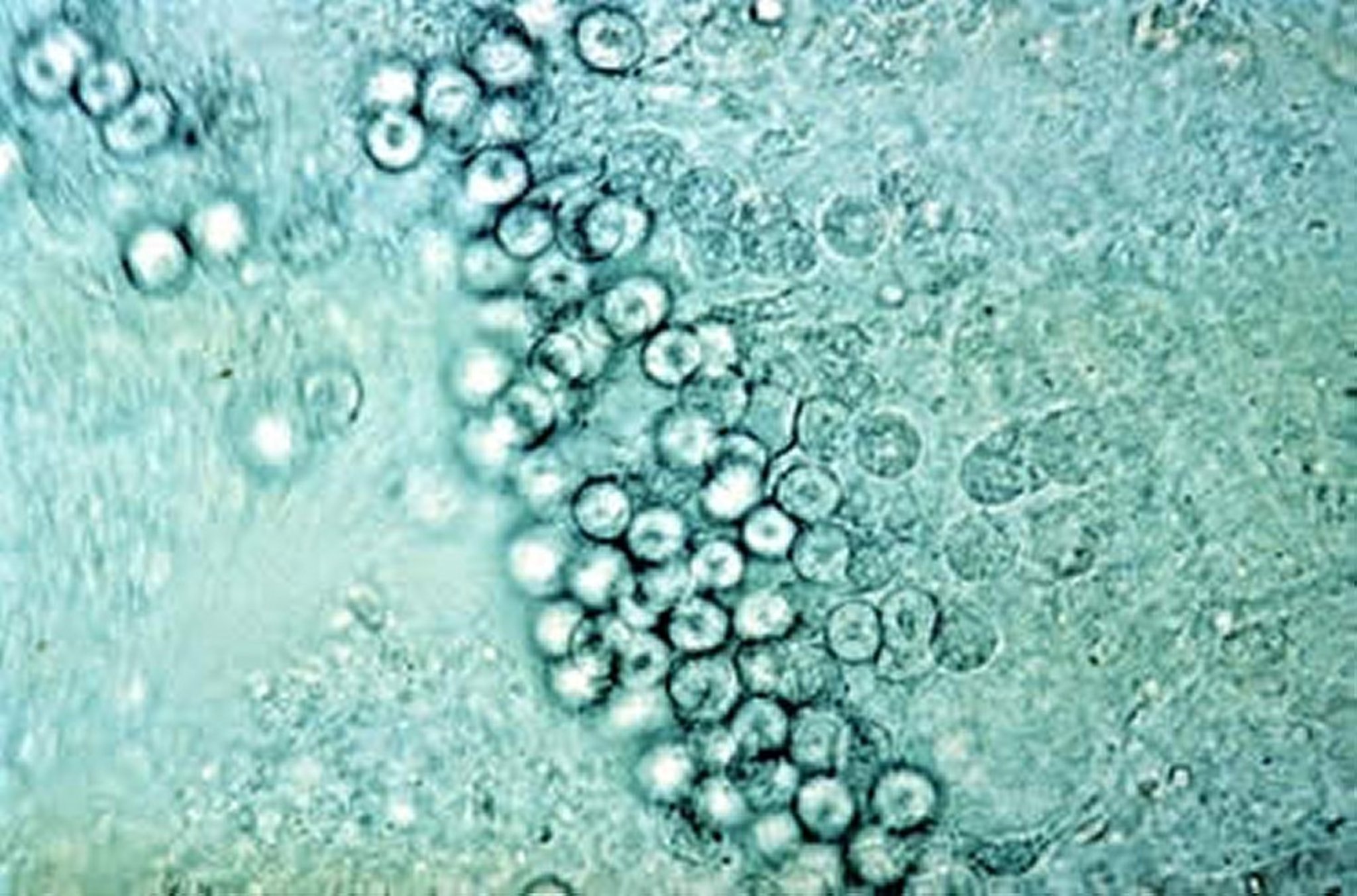
Photomicrograph showing developmental stages of E acervulina from a mucosal scraping of a duodenal loop, poultry. New methylene blue, 40X.
Courtesy of Dr. Jean Sander.

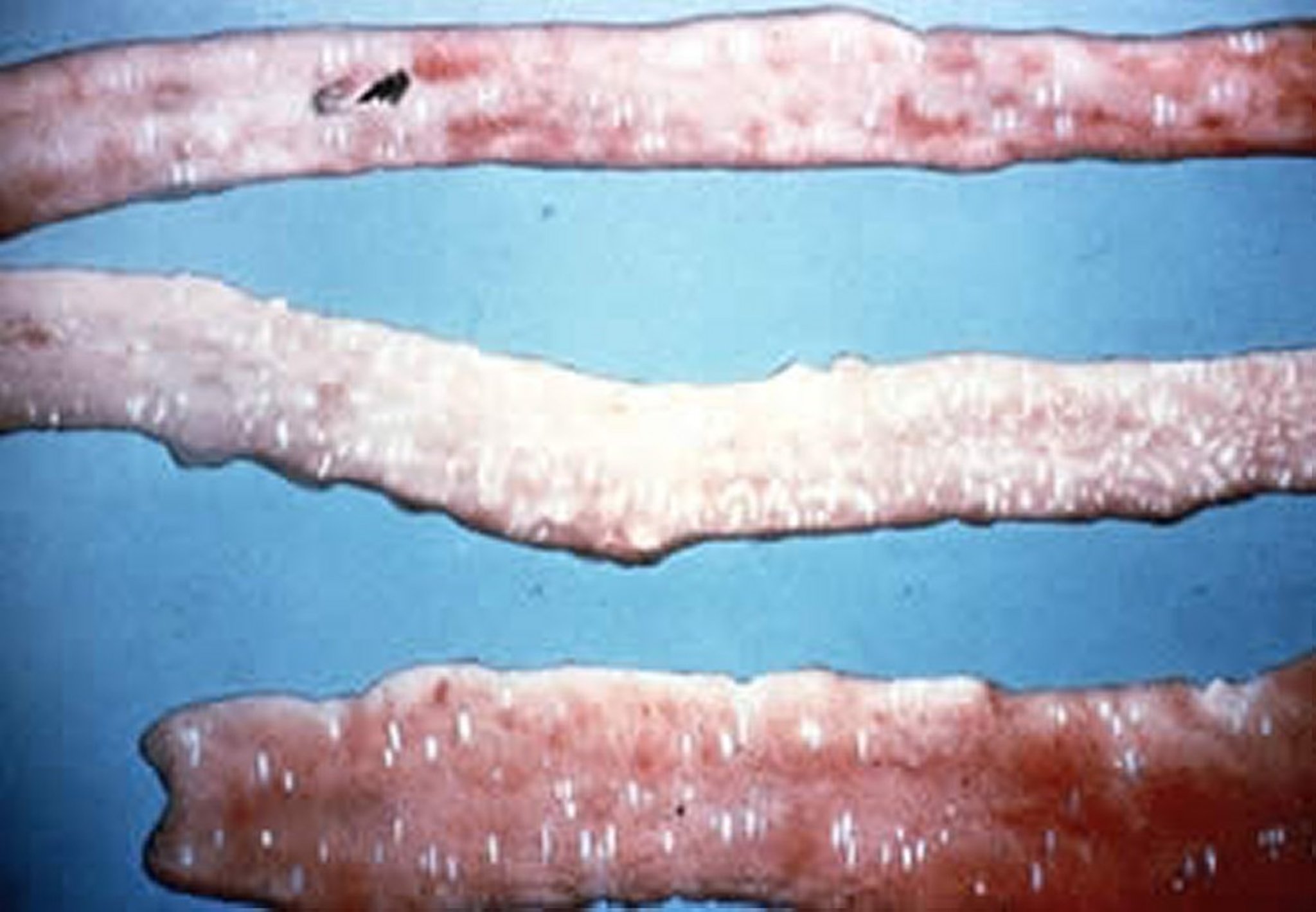
Gross lesions at necropsy in a broiler chicken infected with E acervulina. White longitudinal plaques in the duodenal loop are evident.
Courtesy of Dr. Jean Sander.

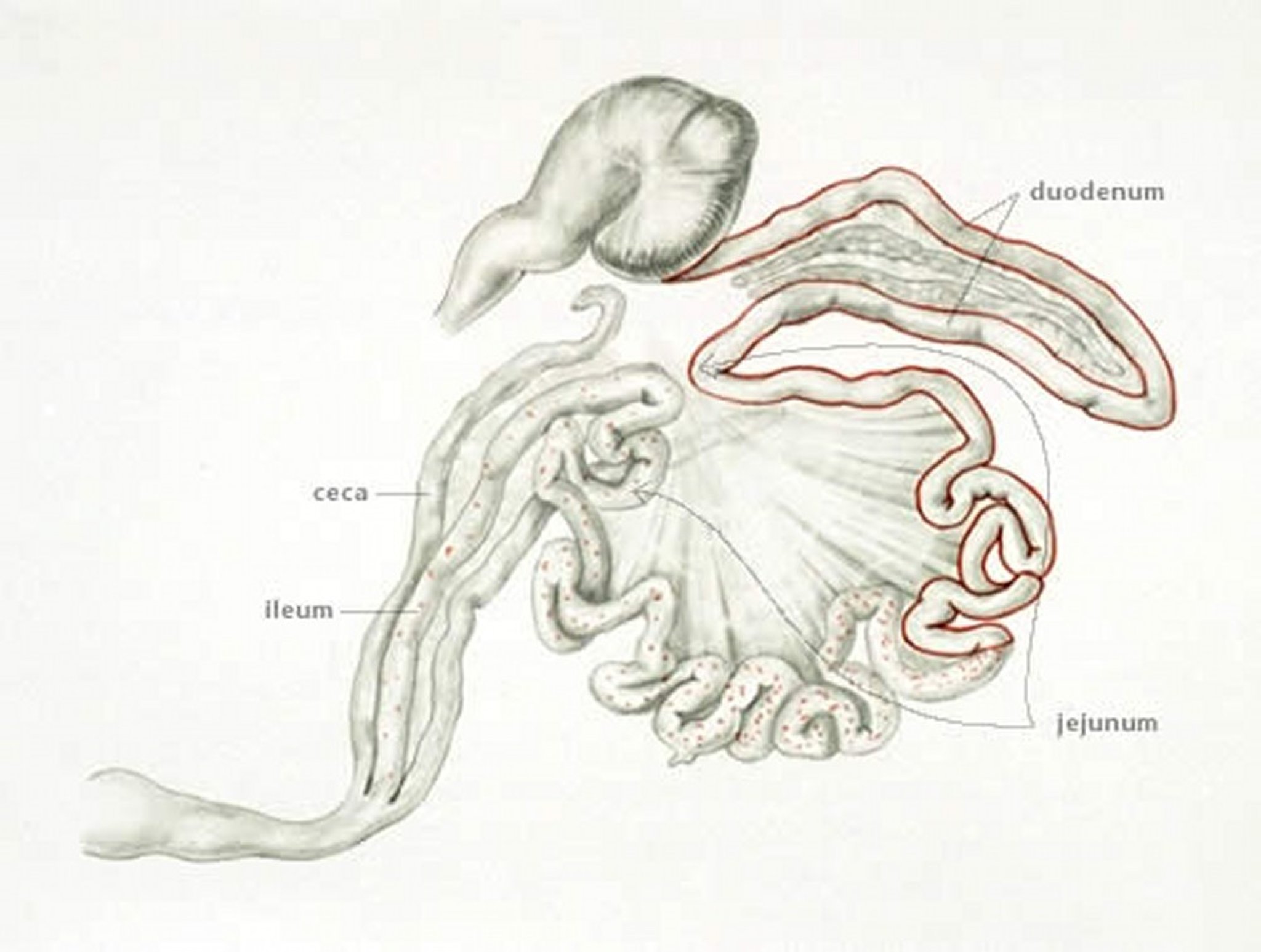
Anatomic drawing illustrating the site of infection with E acervulina in poultry. Characteristic white pinpoint-sized foci in duodenum and jejunum of chickens caused by Eimeria acervulina.
Anatomic drawing by Dr. Gheorghe Constantinescu.
E acervulina is the most common cause of infection. Lesions include numerous whitish, oval or transverse patches in the upper half of the small intestine, which may be easily distinguished on gross examination. The clinical course in a flock is usually protracted and results in poor growth, an increase in culls, and slightly increased mortality.
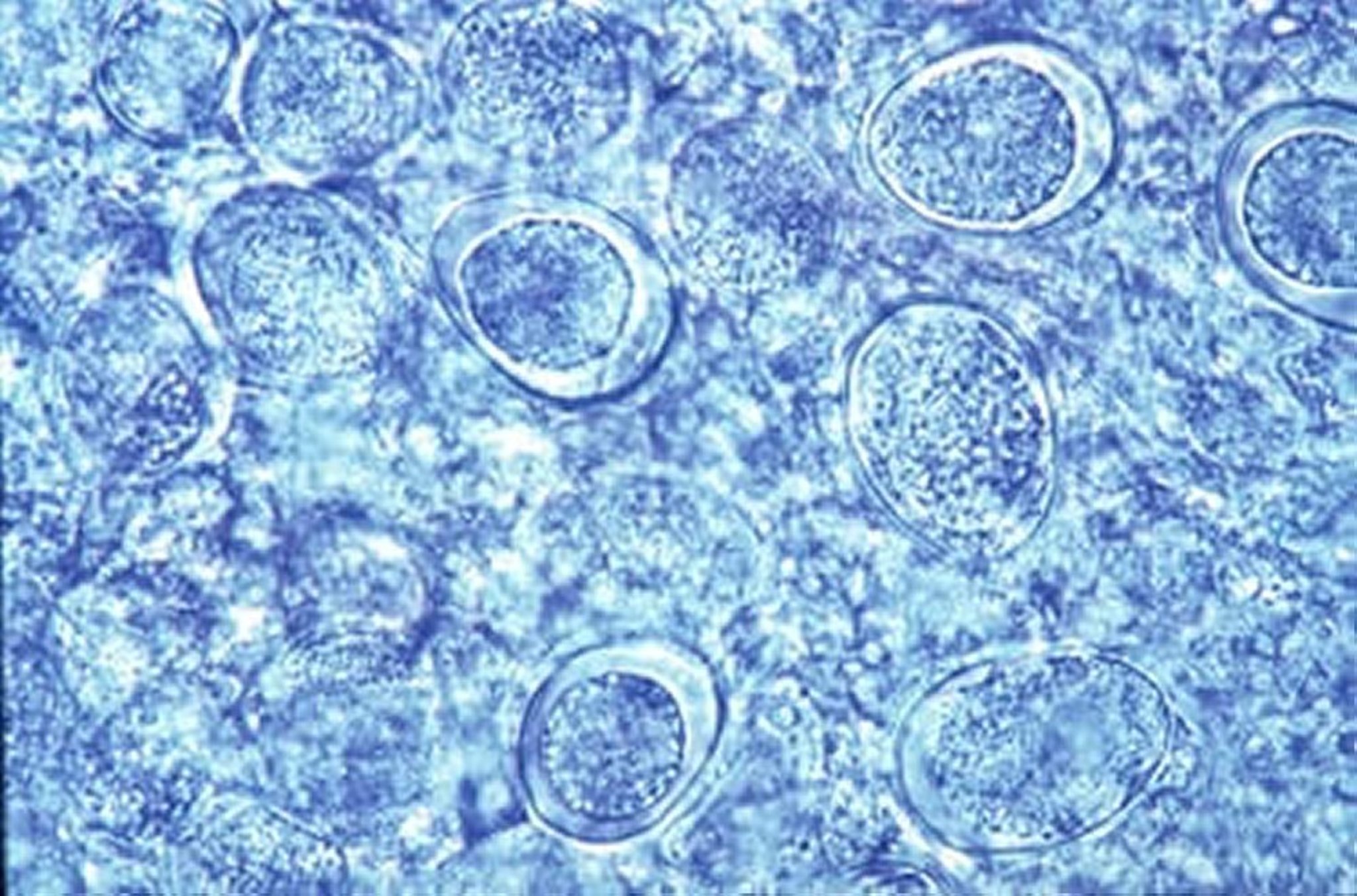
Oocysts of E brunetti from mucosal scraping of small intestine, poultry. New methylene blue, 100X.
Courtesy of Dr. Jean Sander.

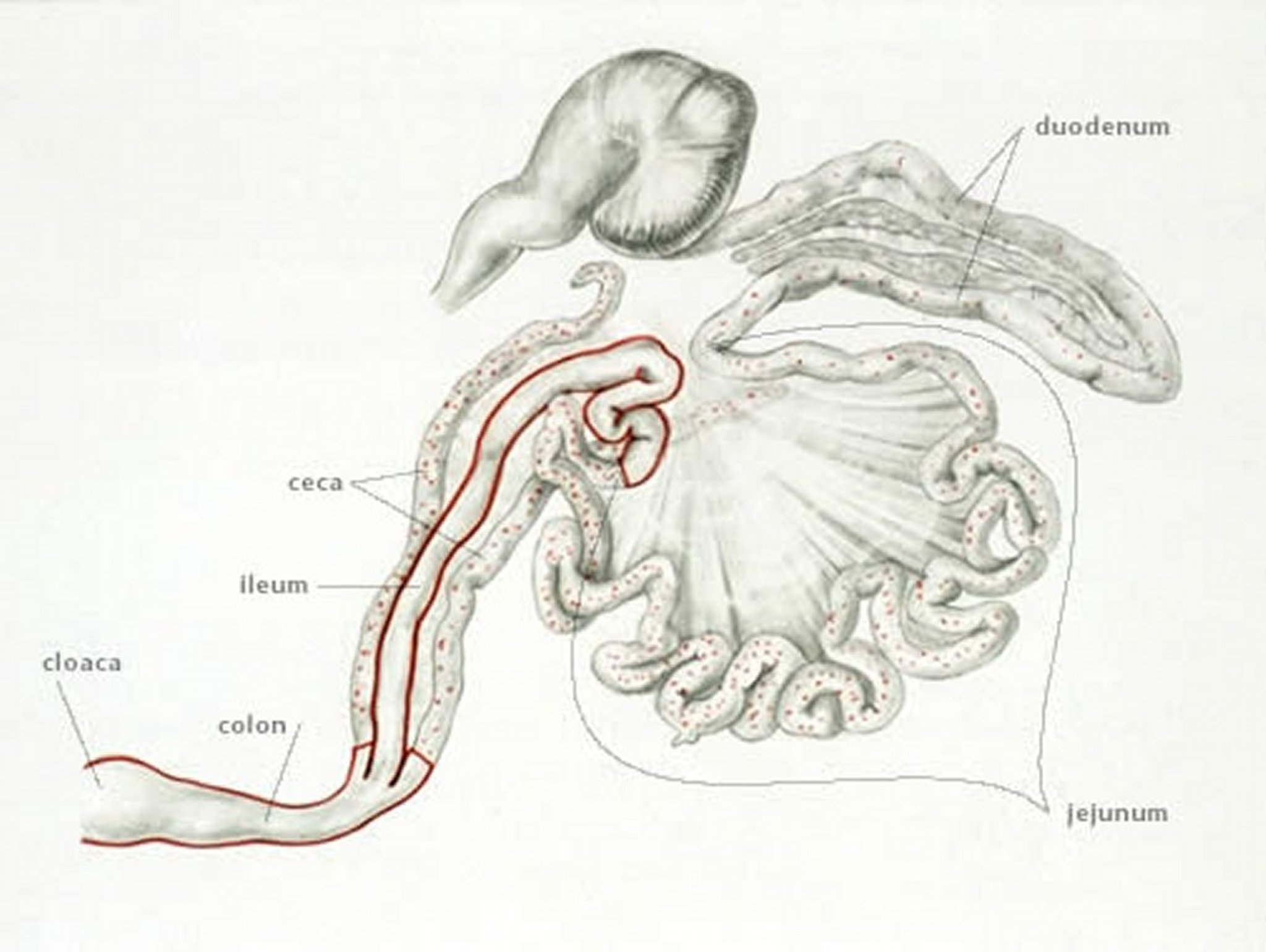
Anatomic drawing of the regions of the chicken gastrointestinal tract illustrating infection with Eimeria brunetti.
Anatomic drawing by Dr. Gheorghe Constantinescu.
E brunetti is found in the lower small intestine, rectum, ceca, and cloaca. In moderate infections, the mucosa is pale and disrupted but lacking in discrete foci, and it may be thickened. In severe infections, coagulative necrosis and sloughing of the mucosa occurs throughout most of the small intestine.

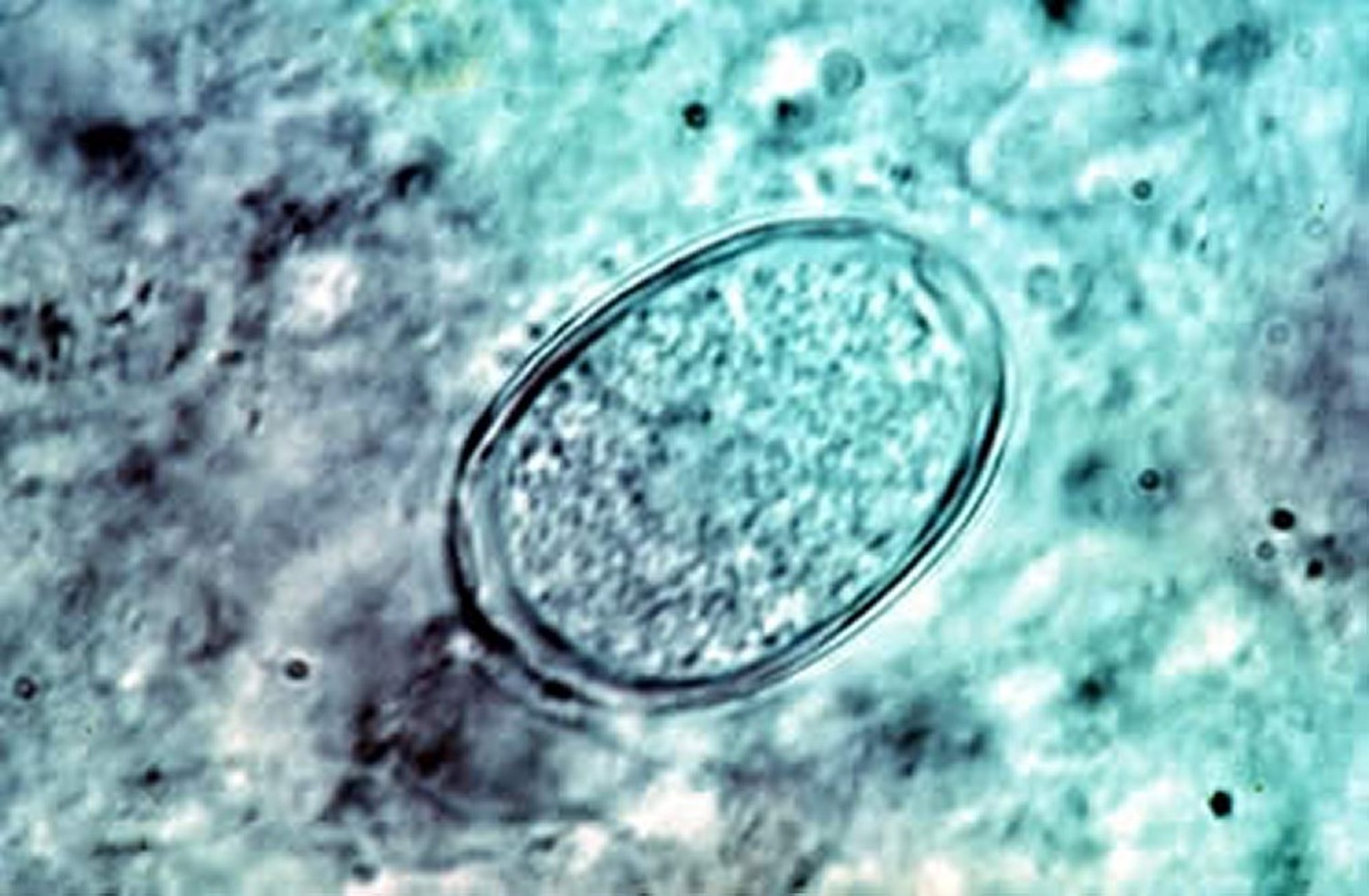
Eimeria maxima oocyst in the feces of an infected chicken.
Courtesy of Dr. Jean Sander.

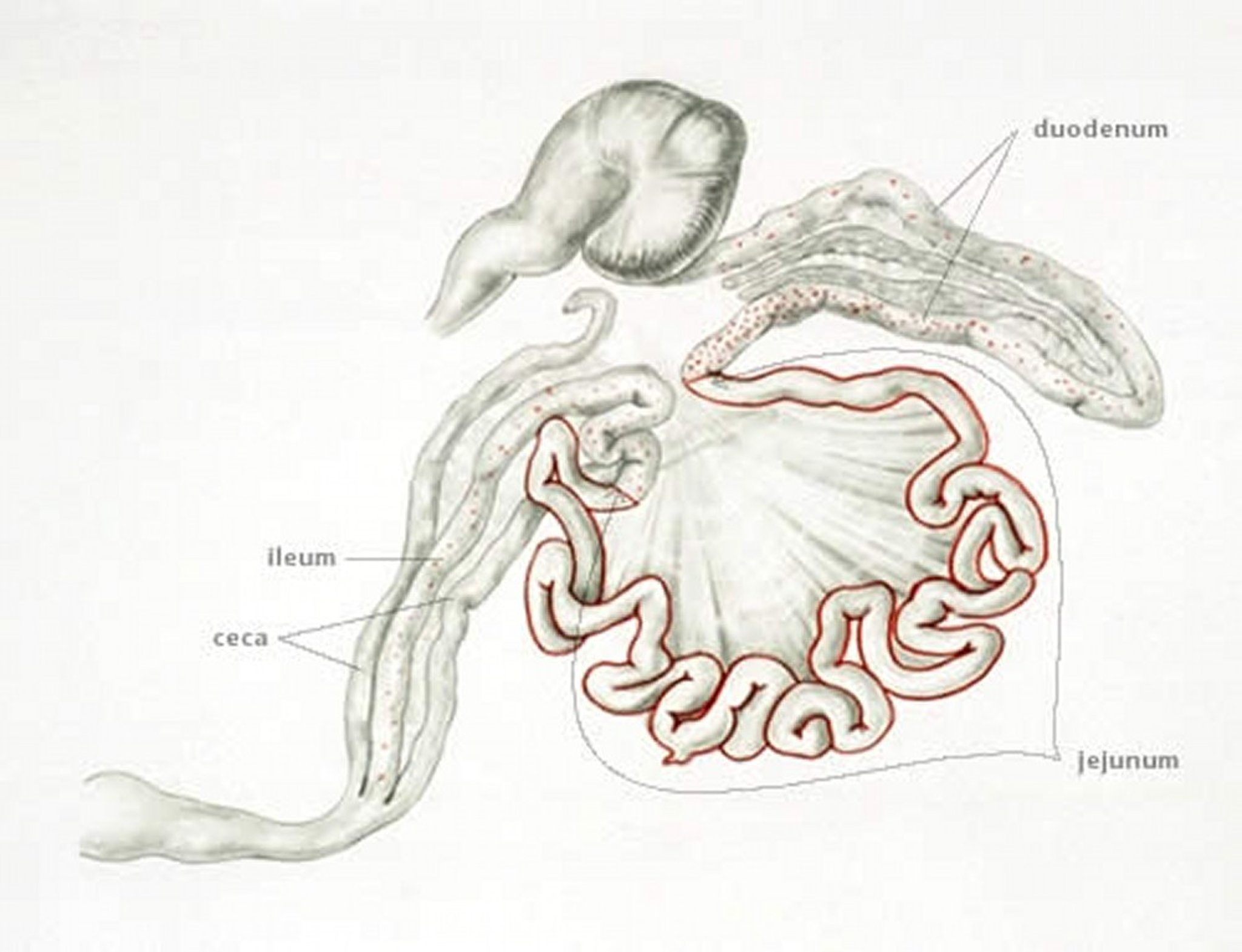
Anatomic drawing illustrating the small intestine as the site of infection with Eimeria maxima in chickens.
Anatomic drawing by Dr. Gheorghe Constantinescu.
E maxima develops in the small intestine, where it causes dilatation and thickening of the wall, petechial hemorrhage, and a reddish orange/pink viscous fluid exudate. The midgut serosa often has numerous whitish pinpoint foci, and may appear engorged. The oocysts and gametocytes (particularly macrogametocytes), present in the lesions, are distinctly large.

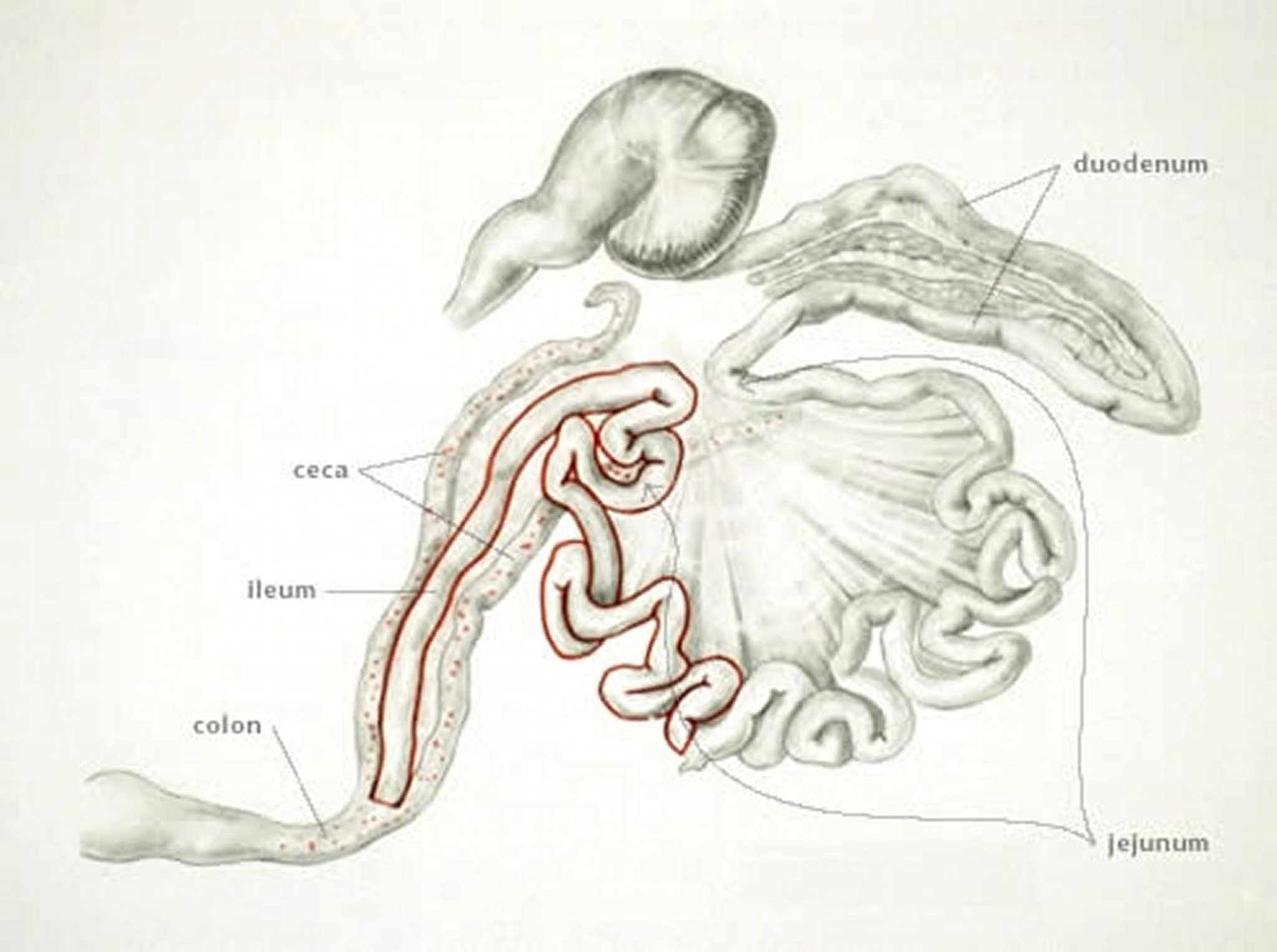
Anatomic drawing illustrating the distal portions of the small intestines as the site of infection with Eimeria mitis in chickens.
Anatomic drawing by Dr. Gheorghe Constantinescu.
E mitis affects the distal small intestine. Lesions are indistinct but may resemble moderate infections of E brunetti. E mitis can be distinguished from E brunetti by finding small, round oocysts associated with the lesion.
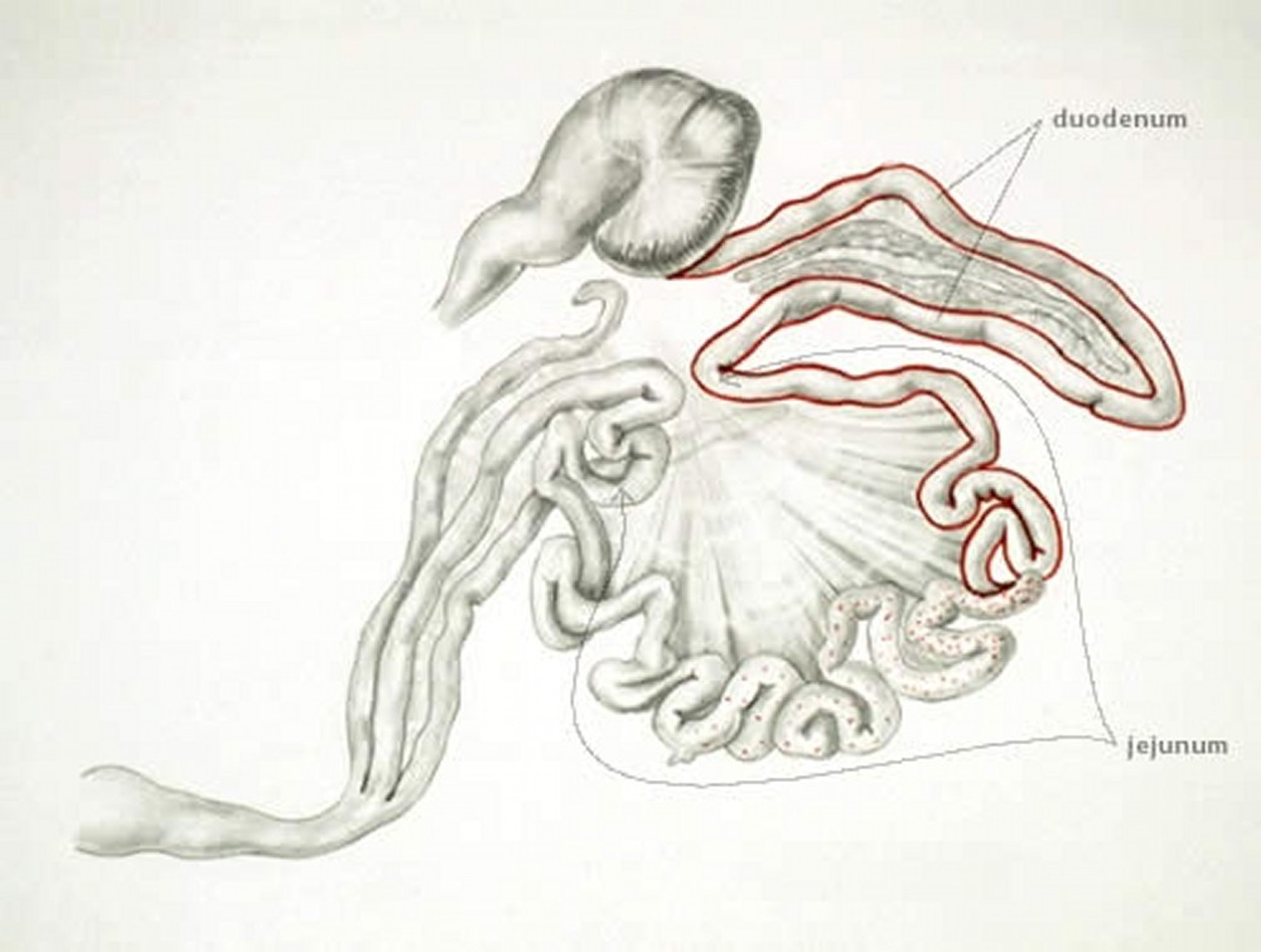
Anatomic drawing illustrating the regions of the proximal small intestine infected with Eimeria praecox.
Anatomic drawing by Dr. Gheorghe Constantinescu.
E praecox, which infects the proximal small intestine, does not cause distinct lesions but may impair growth. The oocysts are larger than those of E acervulina and are numerous in affected areas. The intestinal contents may be watery. E praecox is considered to be of less economic importance versus other species.
E hagani and E mivati develop in the proximal small intestine. The lesions of E hagani are indistinct and difficult to characterize. However, E mivati may cause severe lesions similar to those of E acervulina. It is unclear whether E mivati and E hagani are separate species or variations in size of other known Eimeria spp.
Turkeys
Only four of the seven species of coccidia in turkeys are considered pathogenic: E adenoides, E dispersa, E gallopavonis, and E meleagrimitis. E innocua, E meleagridis, and E subrotunda are considered nonpathogenic. Oocysts sporulate within 1–2 days after expulsion from the host; the prepatent period is 4–6 days.
E adenoeides and E gallopavonis infect the distal ileum, ceca, and rectum. These species often cause death in infected birds. The developmental stages are found in the epithelial cells of the villi and crypts. The affected portion of the intestine may be dilated and have a thickened wall. Thick, creamy material or caseous casts in the gut or feces may contain very large numbers of oocysts. E meleagrimitis mainly infects the proximal and mid small intestine. The lamina propria or deeper tissues may be parasitized, which may result in necrotic enteritis. E dispersa infects the proximal small intestine and causes a creamy, mucoid enteritis that involves the entire intestine, including the ceca. Large numbers of gametocytes and oocysts are associated with the lesions.
Common signs in infected flocks include:
reduced feed consumption
rapid weight loss
lethargy
ruffled feathers
severe diarrhea
watery mucoid feces
Clinical infections are seldom seen in poults > 8 weeks old. Morbidity and mortality may be high.
Game Birds
Popular game birds including the Chinese ringneck pheasant, the chukar partridge, and the bobwhite quail; are reared in large numbers in conditions similar to those for chickens. Losses in these birds due to coccidiosis often exceed 50% of a flock. For common coccidia species in game birds, see see Table: Coccidia Species that Infect Game Birds. Treatment and control are similar to that in poultry; however, amprolium appears to be of little use. Monensin and salinomycin are the approved drugs for quail, and lasalocid and sulfadimethoxine/ormetoprim are the approved drugs for chukars.
Coccidia Species that Infect Game Birds
Game Bird | Coccidia Species |
|---|---|
Pheasant | Eimeria phasiani, E colchici, E duodenalis, E tetartooimia, E pacifica |
Chukar partridge | E kofoidi, E legionensis |
Bobwhite quail | E lettyae, E dispersa, E coloni |
Ducks
A large number of specific coccidia have been reported in both wild and domestic ducks, but validity of some of the descriptions is questionable. Presence of Eimeria, Wenyonella, and Tyzzeria spp has been confirmed. T perniciosa is a known pathogen that causes dilation of the entire small intestine with mucohemorrhagic or caseous material. Eimeria spp also have been described as pathogenic. Some species of coccidia of domestic ducks are considered relatively nonpathogenic. In wild ducks, infrequent but dramatic outbreaks of coccidiosis occur in ducklings 2–4 weeks old; morbidity and mortality may be high.
Geese
The best known coccidial infection of geese is that produced by E truncata, in which necropsy examination shows enlarged kidneys covered with poorly circumscribed, yellowish-white streaks and spots. The tubules are dilated with masses of oocysts and urates. Mortality may be high. At least five other Eimeria spp have been reported to parasitize the intestine of geese, but these are of lesser importance.
Diagnosis
Fecal flotation
Histopathologic examination of tissue samples collected at necropsy
Diagnosis is based on the location of lesions in the host and their appearance; the size of oocysts present is used to determine the species. Coccidial infections are readily confirmed by demonstration of oocysts in feces or intestinal scrapings; however, the number of oocysts present has little relationship to the extent of clinical disease. Severity of lesions as well as knowledge of flock appearance, morbidity, daily mortality, feed intake, growth rate, and rate of lay are important for diagnosis. Necropsy of several fresh specimens is advisable. Classic lesions of E tenella and E necatrix are pathognomonic, but infections with other species are more difficult to diagnose. Comparison of lesions and other clinical signs allows a reasonably accurate differentiation of the coccidial species. Mixed coccidial infections are common.
A diagnosis of clinical coccidiosis is warranted if oocysts, merozoites, or schizonts are seen microscopically and if lesions are severe. Subclinical coccidial infections may be unimportant, and poor performance may be caused by other flock disorders.
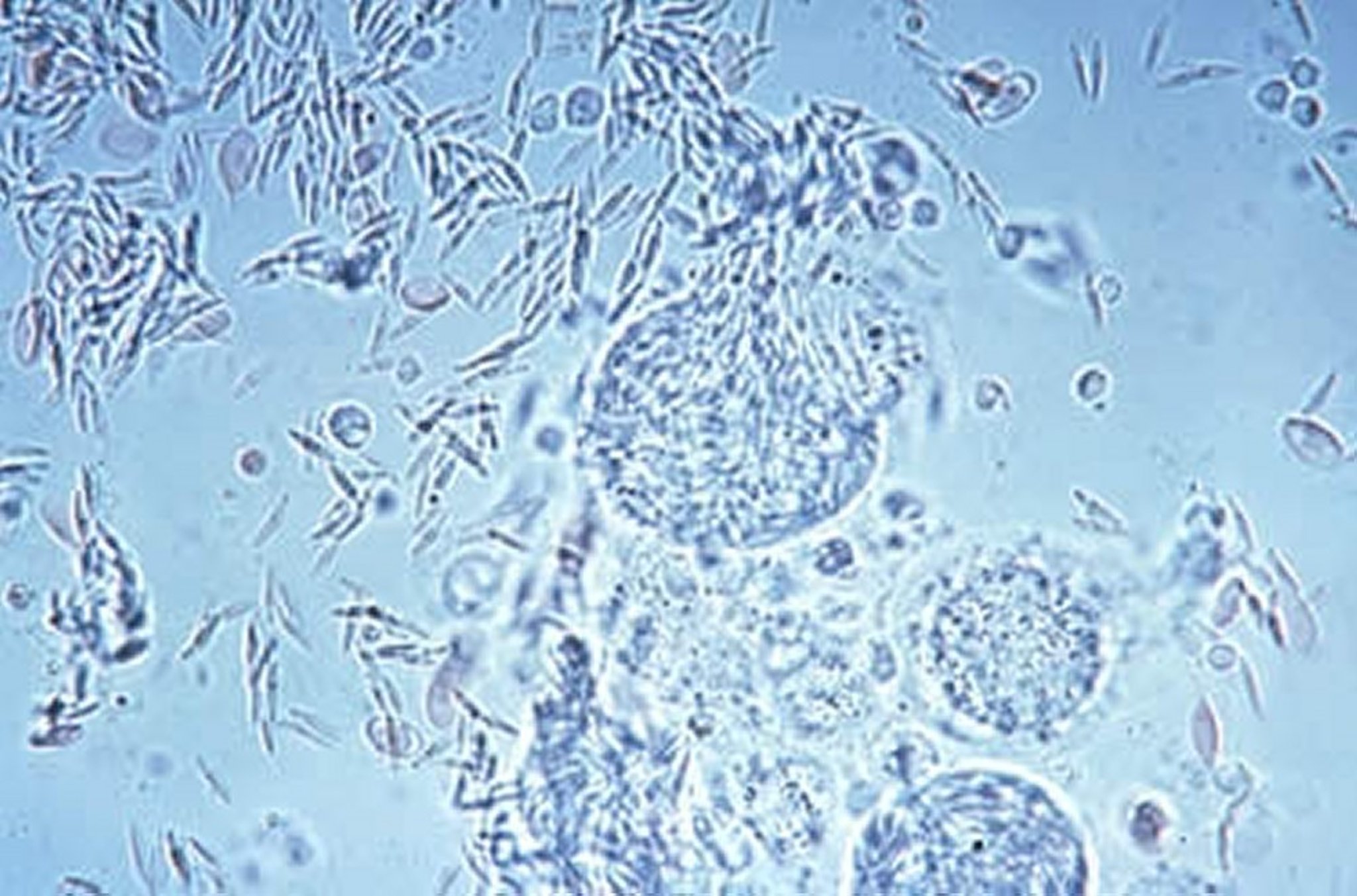
Merozoites being released from intracellular schizonts. Released merozoites can infect another host intestinal cell, continuing the asexual replication of the parasite.
Courtesy of Dr. Jean Sander.
Control
Use of anticoccidial compounds in feed or water, vaccination, or a combination of both to prevent clinical signs
Once clinical signs appear, use of antibiotics and supportive care is advisable to minimize dehydration and secondary bacterial infection
Practical methods of management cannot prevent coccidial infection. Poultry maintained at all times on wire floors to separate birds from feces have fewer infections; clinical coccidiosis is seen only rarely under such circumstances. Other methods of control are vaccination or prevention with anticoccidial drugs.
Vaccination
A species-specific immunity develops after natural infection, the extent largely depends on the severity of infection and the number of reinfections. Protective immunity is primarily a T-cell response.
Commercial vaccines consist of live, sporulated oocysts of the various coccidial species administered at low doses. Modern anticoccidial vaccines should be given to day-old chicks, either at the hatchery or on the farm. Because the vaccine serves only to introduce infection, chickens are reinfected by progeny of the vaccine strain on the farm. Most commercial vaccines contain live oocysts of coccidia that are not attenuated. The self-limiting nature of coccidiosis is used as a form of attenuation for some vaccines rather than biologic attenuation. Some vaccines sold in Europe and South America include attenuated lines of coccidia. Research has shown promise for vaccination in game birds.
Layers and breeders maintained on floor litter must have protective immunity. Historically, these birds were given a suboptimal dosage of an anticoccidial drug during early growth, with the expectation that immunity would continue to develop from repeated exposure to wild types of coccidia. This method has never been completely successful because of the difficulty in controlling all the factors affecting reproduction of coccidia under practical conditions. Although anticoccidial drugs have been preferred for protection of these birds, vaccination programs are gaining popularity. More effective administration techniques and choice of coccidia strains in the product are improving the feasibility of vaccination in broilers.
Anticoccidial Drugs
Many products are available for prevention or treatment of coccidiosis in chickens and turkeys. Detailed instructions for use are provided by all manufacturers to help users with management considerations and to ensure compliance with regulatory requirements.
Anticoccidials are given in the feed to prevent disease and the economic loss often associated with subacute infection. Prophylactic use is preferred, because most of the damage occurs before signs become apparent and because drugs cannot completely stop an outbreak. Therapeutic treatments are usually given by water because of the logistical constraints on feed administration. Antibiotics and increased levels of vitamins A and K are sometimes used in the ration to improve rate of recovery and prevent secondary infections.
Continuous use of anticoccidial drugs promotes the emergence of drug-resistant strains of coccidia. Various programs are used in attempts to slow or stop selection of resistance. For instance, producers may use one anticoccidial continuously through successive flocks, change to alternative anticoccidials every 4–6 months, or change anticoccidials during a single growout (ie, a shuttle program). Although there is little cross-resistance to anticoccidials with different modes of action, there is widespread resistance to most drugs.
Coccidia can be tested in the laboratory to determine which products are most effective. “Shuttle programs,” in which one group of chickens is treated sequentially with different drugs (usually a change between the starter and grower rations), are common practice and offer some benefit in slowing the emergence of resistance. Shuttle programs are considered extralabel use in some jurisdictions (eg, the US) and should be used only as permitted by law.
The effects of anticoccidial drugs may be coccidiostatic, in which growth of intracellular coccidia is arrested but development may continue after drug withdrawal, or coccidiocidal, in which coccidia are killed during their development. Some anticoccidial drugs may be coccidiostatic when given short-term but coccidiocidal when given longterm. Most anticoccidials currently used in poultry production are coccidiocidal.
The natural development of immunity to coccidiosis may proceed during the use of anticoccidials in the feed. However, in the production of broilers during a short growout of 37–44 days, this may be of little consequence. Natural immunity is important in replacement layer pullets, because they are likely to be exposed to coccidial infections for extended periods after termination of anticoccidial drugs. Anticoccidial programs for layer and breeder flocks are intended to allow immunizing infection while guarding against acute outbreaks.
Anticoccidials are commonly withdrawn from broilers 3–7 days before slaughter to meet regulatory requirements and to reduce production costs. Because broilers have varying susceptibility to infection at this point, the risk of coccidiosis outbreaks is increased with longer withdrawal.
Turkeys are given a preventive anticoccidial for confinement-reared birds up to 8–10 weeks of age. Older birds are considered less susceptible to outbreaks.
The modes of action of anticoccidial drugs are poorly understood. Knowledge of mode of action is important in understanding potential adverse effects.
Amprolium is an antagonist of thiamine (vitamin B1). Rapidly dividing coccidia have a high requirement for thiamine. Amprolium has a safety margin of ~8:1 when used at the highest recommended level in feed (125–250 ppm). Because amprolium has poor activity against some Eimeria spp, its spectrum has been extended by using it in mixtures with the folic acid antagonists ethopabate and sulfadimethoxine. The primary use of amprolium currently is for water treatment during clinical outbreaks.
Clopidol (eg, decoquinate) is coccidiostatic against early development of Eimeria spp by inhibiting mitochondrial energy production. Clopidol has a broad species spectrum. However, resistance may develop rapidly during extended use.
Folic acid antagonists include the sulfonamides (not all legally approved) and ethopabate. These compounds are structural antagonists of folic acid or of para-aminobenzoic acid (PABA), which is a precursor of folic acid. (The host does not synthesize folic acid and has no requirement for PABA.) Coccidia rapidly synthesize nucleic acids, accounting for activity of PABA antagonists. Although resistance to antifolate compounds is widespread, they are commonly used for water treatment when clinical signs are already evident. Ormetoprim is active against the protozoan enzyme dihydrofolate reductase. It has synergistic activity with sulfonamides and often is used in mixtures with these compounds.
Halofuginone hydrobromide is related to the antimalarial drug febrifuginone and is effective against asexual stages of most species of Eimeria. It has both coccidiostatic and coccidiocidal effects, but coccidia may become resistant after extended exposure.
The ionophores (monensin, salinomycin, lasalocid, narasin, and maduramicin) form complexes with various ions, principally sodium, potassium, and calcium, and transport these into and through biologic membranes. The ionophores affect both extra- and intracellular stages of the parasite, especially during the early, asexual stages of parasite development. Drug tolerance was slow to emerge in chicken coccidia, probably because of the biochemically nonspecific way these fermentation products act on the parasite. Recent surveys suggest that drug tolerance is now widespread, but these products remain the most important class of anticoccidials. Farms claiming to be antibiotic-free cannot use ionophores because they are classified as antibiotics in the US.
Some ionophores may depress feed consumption when the dosage is above recommended levels. Primarily, this is the result of reduced feed consumption, but the reduced growth may be offset by improved feed conversion.
Nicarbazin was the first product to have truly broad-spectrum activity and has been in common use since 1955. Although not completely understood, the mode of action is thought to be via inhibition of succinate-linked nicotinamide adenine dinucleotide reduction and the energy-dependent transhydrogenase, and the accumulation of calcium in the presence of ATP. Nicarbazin is toxic for layers, causing mottling of egg yolks, decreased egg production, and blanching of brown egg shells. A 4-day withdrawal period is required in broilers. Medicated birds are at increased risk of heat stress in hot weather.
Robenidine, a guanidine compound, allows initial intracellular development of coccidia but prevents formation of mature schizonts. It is coccidiostatic when given short term and coccidiocidal long term. Drug resistance may develop during use. A 5-day withdrawal period is needed to eliminate untoward flavor caused by residues in poultry meat.
Diclazuril is highly effective against a broad spectrum of coccidia. It is used mostly for prevention at 1 ppm in the feed.
Key Points
Coccidiosis is a disease of young animals and is characterized by diarrhea and weight loss.
Coccidia are species specific, so cross infection is not seen.
Diagnosis is via fecal flotation or postmortem examination of tissues of affected birds.
Prevention of the disease is key, using various anticoccidial drugs and/or vaccination.


