

Infectious bursal disease is seen in young domestic chickens worldwide and is caused by infectious bursal disease virus (IBDV). Clinical signs include listlessness, watery diarrhea, ruffled feathers, and dehydration. Morbidity rate is high and mortality rate is usually low, but some virulent strains cause mortality rates of 60% or higher. Macroscopic and microscopic lesions in the cloacal bursa and molecular identification of the viral genome are used for diagnosis. Vectored and attenuated live virus vaccines can be used to induce active immunity in chicks as the maternal antibodies wane.
Infectious bursal disease is an economically important viral disease of young domestic chickens worldwide. Infectious bursal disease was first identified in Gumboro, Delaware, in 1962.
Etiology of Infectious Bursal Disease
The etiologic agent of infectious bursal disease is the infectious bursal disease virus (IBDV), a double-stranded RNA virus (Avibirnavirus gumboroense) in the family Birnaviridae. As with other birnaviruses, IBDV virions are nonenveloped and have an icosahedral single-shelled geometry.
Two serotypes of IBDV have been identified. The serotype 1 viruses cause disease in chickens and, within them, antigenic variation can exist between strains. Antigenic drift is largely responsible for this antigenic variation; however, antigenic differences can also occur through genome homologous recombination.
Virulence of serotype 1 field strains varies considerably. Virus strains can be classified by phenotype into subclinical, classical virulent, or very virulent (vv). Alternatively, they can be classified based on the antigenicity of the hypervariable segment of viral protein 2 (VP2) into classical or variant groups.
The cloacal bursa is a primary lymphoid organ located dorsally to the cloaca and is responsible for the development and maturation of B cells in young poultry. Atrophy of the bursa, which includes the loss of B lymphocytes, occurs ~7–10 days after IBDV infection.
Immunosuppression is directly related to this loss of B lymphocytes; however, immunosuppression and related secondary infections are typically seen in birds that recover from the disease. Severity of the immunosuppression depends on the virulence of the infecting virus and age of the host.
Epidemiology of Infectious Bursal Disease
Infectious bursal disease affects young domestic chickens worldwide.
Classical strains predominated until the emergence of variant IBDV strains in 1986. The vvIBDV strains were first detected in Europe in 1989 and spread throughout the Middle East, Asia, and Africa. They were detected in South and Central America in 1999 and in the US in 2009.
Infectious bursal disease is highly contagious. IBDV is shed in the feces and transferred from house to house by fomites.
The variant IBDV strains typically cause subclinical infection in chickens; however, low mortality rates (< 10%) may be observed in some breeds of layer chickens. The classical IBDV strains can cause clinical signs of disease and moderate mortality rates (< 40%), while the vvIBDV strains can cause high mortality rates (> 60%).
Flock morbidity rate is typically 100%, and mortality rate can range from 5% to greater than 60% depending on the strain of virus and breed of chicken. Mortality rate is typically higher in layer breeds compared to broiler chickens.
Chickens are most susceptible to clinical disease at 3–6 weeks, when immature B cells populate the bursa and maternal immunity has waned. However, severe infections have occurred in Leghorn chickens up to 18 weeks old.
In clinical infections, onset of the disease occurs after an incubation period of 3–4 days.
Serotype 2 strains of the virus infect chickens and turkeys but have not caused clinical signs of disease or immunosuppression in these hosts. IBDVs have been identified in other avian species, including penguins, and antibodies against IBDV have been seen in several wild avian species. The contribution of IBDV to disease in these wild birds is unknown.
Clinical Findings of Infectious Bursal Disease
Depending on the IBDV strain and presence of maternal immunity, infectious bursal disease can cause clinical or subclinical disease in young chicks. Results of infection depend on age and breed of chicken and virulence of the virus. Infections before 3 weeks old are usually subclinical.
For both clinical and subclinical forms of the disease, all pathogenic IBDVs cause lesions in the cloacal bursa (bursa of Fabricius).
Early subclinical infections are the most important form of the disease because of economic losses. They cause severe, long-lasting immunosuppression due to destruction of immature lymphocytes in the cloacal bursa, thymus, and spleen.
The humoral (B cell) immune response is most severely affected; the cell-mediated (T cell) immune response is affected to a lesser extent.
Chickens immunosuppressed by early IBDV infections do not respond well to vaccination and are predisposed to infections with normally nonpathogenic viruses and bacteria. Common diseases are usually exacerbated by IBDV infections.
Some strains of IBDV can cause subclinical infections in older birds (3–6 weeks old), leading to losses from poor feed efficiency and longer times to market. In these cases, immunosuppression is usually transient, and convalescent birds may recover most or all of their humoral immune function. However, secondary infections that occur during the transient immunosuppression can cause substantial economic losses.
Following an incubation period of 3–4 days, clinical signs may include:
severe prostration
incoordination
watery diarrhea
soiled vent feathers
vent picking
inflammation of the cloaca
Recovery occurs in < 1 week, and broiler weight gain is delayed by 3–5 days. The presence of maternal antibody will modify the clinical course of the disease.
Lesions

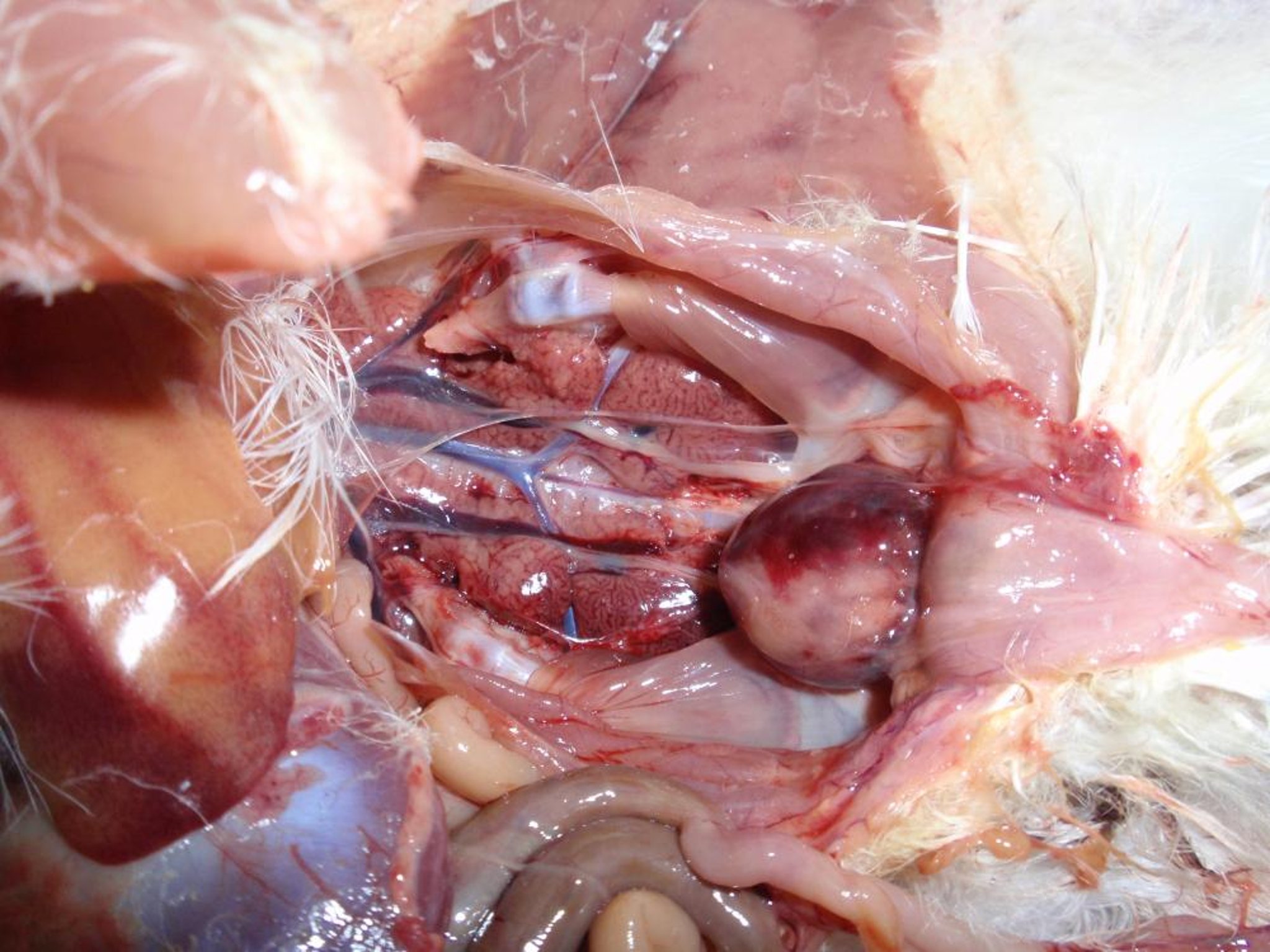
Gross pathology photograph showing an enlarged, hemorrhagic cloacal bursa in a chicken infected with very virulent infectious bursal disease virus. This strain of the virus can also cause hemorrhages in skeletal muscles.
Courtesy of Dr. Daral J. Jackwood.

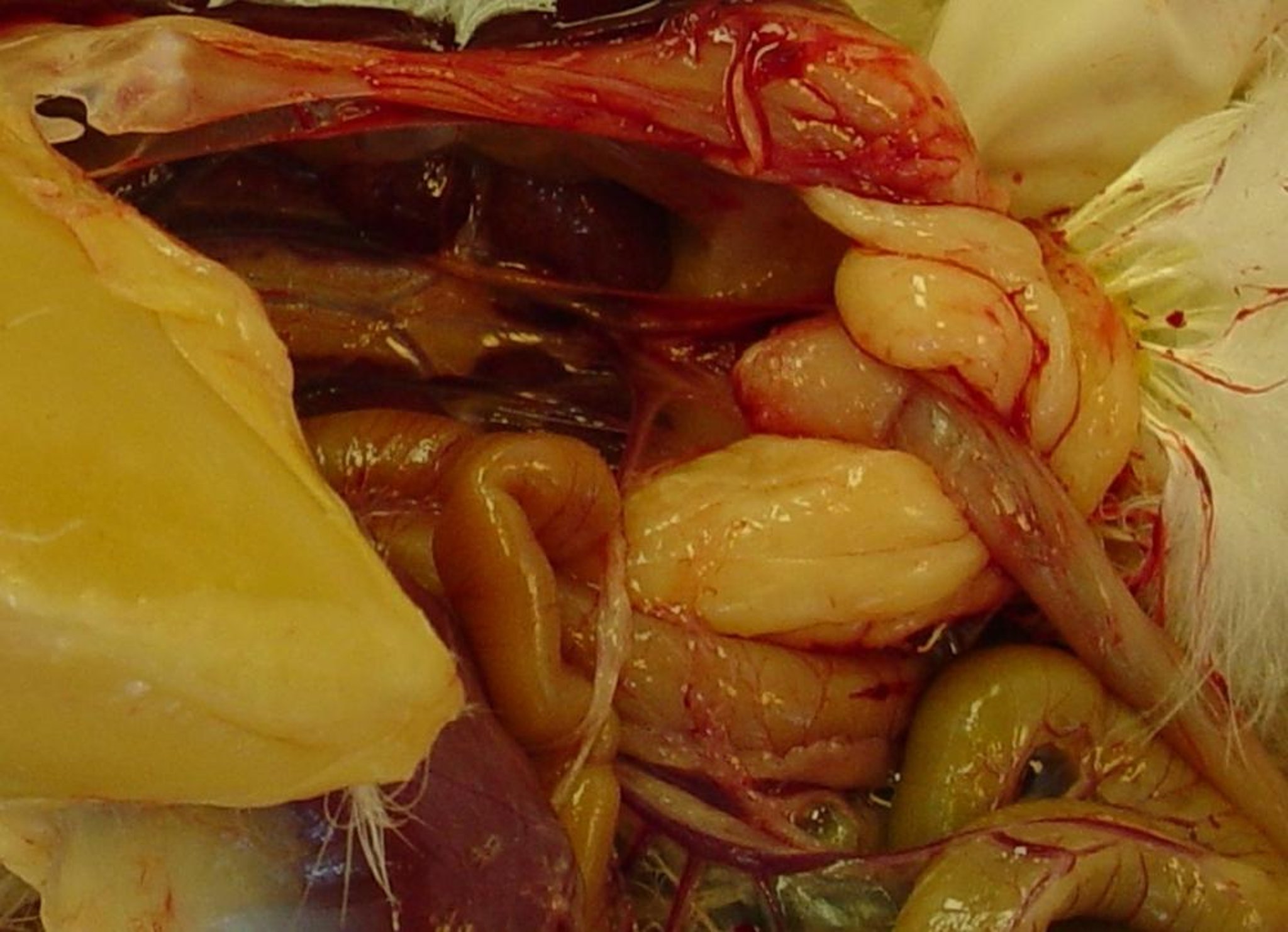
Gross pathology photograph showing a small, atrophic cloacal bursa (albeit without gross lesions) in a subclinically affected chicken infected with a variant strain of infectious bursal disease virus.
Courtesy of Dr. Daral J. Jackwood.
At necropsy, the lesions observed will depend on the strain of IBDV.
For strains that cause signs of disease, the cloacal bursa becomes edematous, with a yellowish-colored transudate on the surface. Hemorrhages on the serosal and mucosal surfaces are sometimes observed.
Strains of vvIBDV cause similar cloacal bursa lesions, and congestion and hemorrhage of the pectoral and leg muscles can also occur. Some IBDV strains can cause atrophy of the cloacal bursa without the appearance of gross lesions in that organ.
Chickens that have recovered from IBDV infections have small, atrophied cloacal bursas due to the destruction and lack of regeneration of the bursal follicles.


A chicken demonstrating severe listlessness, ruffled feathers, and a crouched stance 4 days after infection with IBDV.
Courtesy of Dr. Julia R. Blakey.
Diagnosis of Infectious Bursal Disease
Detection of macroscopic and microscopic lesions in the cloacal bursa
Detection of IBDV viral RNA
Virus isolation
Initial diagnosis of infectious bursal disease is accomplished by the observation of gross lesions in the cloacal bursa. This is followed by microscopic analysis of the bursa for lymphocyte depletion in the follicles.
Molecular diagnostic assays are most often used to identify IBDV in diagnostic samples. IBDV is most readily isolated from the cloacal bursa but may be isolated from other organs. RT-PCR assay is used to identify the viral genome in cloacal bursa tissue.
Sequence alignments and phylogenetic analysis of the VP2 coding region has been used to further characterize the viruses into genogroups. Samples for molecular diagnostic testing are typically collected after maternal antibodies have waned.
IBDV may be isolated in 8- to 11-day-old IBDV antibody–free chicken embryos with inocula from birds in the early stages of disease. The chorioallantoic membrane is more sensitive to inoculation than is the allantoic sac.
Some strains of IBDV may also be isolated in cell cultures that include chicken embryo fibroblasts, cells from the cloacal bursa, and established avian and mammalian cell lines. Cell culture–adapted strains of IBDV produce a cytopathic effect and may be used for quantitative titration of the virus and virus-neutralization assays.
Serologic testing can be used to detect antibodies against IBDV in convalescent chicks. Commercially available ELISA kits are most often used to quantitate IBDV antibodies. The presence of IBDV antibodies in chicks is not always an indication of infection because most young chicks have maternal antibodies.
Differential diagnoses for cloacal bursal atrophy include:
IBDV vaccination
stress
In addition, normal bursal involution can be confused with the atrophic cloacal bursa. The cloacal bursa reaches peak size at 6 weeks and then undergoes involution over a period of several months, with complete involution occurring at sexual maturity at 16–24 weeks.
Treatment and Control of Infectious Bursal Disease
There is no treatment for infectious bursal disease. Therefore, control and prevention are key.
Rigorous disinfection of contaminated farms after depopulation has achieved limited success. IBDV is very stable in the environment and difficult to eradicate from premises.
Live, attenuated virus vaccines of chicken embryo or cell-culture origin and of varying low pathogenicity can be administered by eye drop, drinking water, or SC routes at 1–21 days old. Replication of these vaccines and thus the immune response can be altered by maternal antibody, although the more virulent vaccine strains can override higher concentrations of maternal antibody.
Vectored vaccines that express the IBDV VP2 protein in herpesvirus of turkeys (HVT) can be administered in ovo or at hatch. These vaccines are not affected by maternal antibodies. Vaccines that use live, attenuated viruses bound to antibodies (immune-complex vaccines) are also available for in ovo or at hatch administration.
High concentrations of maternal antibody during early brooding of chicks in broiler flocks (and in some commercial layer operations) can minimize early infection, subsequent immunosuppression, or both.
Breeder flocks should be vaccinated one or more times during the growing period, first with a live, attenuated virus vaccine and again just before egg production with an oil-adjuvanted, inactivated vaccine.
Inactivated vaccines of chicken embryo, bursa, or cell-culture origin are available. The latter vaccines induce higher, more uniform, and more persistent concentrations of antibody than do live, attenuated virus vaccines.
The immune status of breeder flocks should be monitored periodically with a quantitative serologic test such as virus neutralization or ELISA. If antibody concentrations decrease, hens should be revaccinated to maintain adequate immunity in the progeny.
Vaccination programs should aim to use vaccines that most closely match the antigenic profile of the field viruses. Diagnostic testing for the genomic sequences of field strains can be used to select the most appropriate vaccination program.
Key Points
Infectious bursal disease, caused by infectious bursal disease virus, is a disease of young chickens found worldwide.
IBDV infects immature B lymphocytes and causes an immune suppression that leads to secondary infections in convalescent birds.
Diagnosis is through clinical evaluation of the cloacal bursa and molecular identification of the viral genome.
Control is accomplished using vaccination of breeder flocks to induce maternal immunity in young chicks.


