



Marek's disease is a highly contagious viral disease of poultry characterized by T-cell lymphomas and peripheral nerve enlargement. Standard criteria used for diagnosis include history, clinical signs, gross necropsy, and histological examination. Although no treatment is available, current vaccines are highly protective.
Chickens are the most important natural host for Marek's disease virus (MDV), a highly cell-associated but readily transmitted alphaherpesvirus with lymphotropic properties of gammaherpesviruses. Quail and turkeys can also be infected, especially when they are raised in proximity to chicken farms.
Turkeys are also commonly infected with turkey herpesvirus (HVT), an avirulent strain related to Marek's disease virus that is commonly used as a Marek's disease vaccine in chickens. Although Marek's disease has been reported in other birds, such as pheasants and peafowl, mammals appear to be refractory to the infection.
Marek's disease is one of the most ubiquitous avian infections; it is identified in chicken flocks worldwide. Every flock, except for those maintained under strict pathogen-free conditions, is presumed to be infected. Although clinical signs of disease are not always apparent in infected flocks, a subclinical decrease in growth rate and egg production may be economically important.
Etiology of Marek's Disease in Poultry
Marek's disease virus is a member of the genus Mardivirus within the subfamily Alphaherpesvirinae. Within the genus Mardivirus are three closely related species previously designated as three serotypes of Marek's disease virus:
Gallid alphaherpesvirus 2 (MDV serotype 1) represents all virulent Marek's disease virus strains and is further divided into pathotypes, designated as mild (m), virulent (v), very virulent (vv), and very virulent plus (vv+).
Gallid alphaherpesvirus 3 (MDV serotype 2) and Meleagrid alphaherpesvirus 1 (turkey herpesvirus, MDV serotype 3) represent avirulent virus strains isolated from chickens and turkeys, respectively, and are commonly used as vaccines against Marek's disease.
Pathogenesis of Marek's Disease in Poultry
Currently, four phases of infection with Marek's disease are recognized:
Early cytolytic infection (productive-restrictive)
Latent infection
Second phase of cytolytic, productive-restrictive infection coincident with permanent immunosuppression
Proliferative phase, involving nonproductively infected lymphoid cells that may or may not progress to the point of lymphoma formation
Productive infection may occur transiently in B lymphocytes within a few days after infection with virulent Marek's disease virus strains and is characterized by antigen production, which leads to cell death. Because few if any virions are produced, this has also been termed a restrictive-productive infection. Productive infection also occurs in the feather follicle epithelium, in which enveloped virions are produced.
Latent infection of activated T cells is responsible for the longterm carrier state. No antigens are expressed, but virus can be recovered from such lymphocytes by cocultivation with susceptible cells in tissue cultures. Some T cells, latently infected with oncogenic MDV strains, undergo neoplastic transformation. These transformed cells may multiply to form characteristic lymphoid neoplasms.
Epidemiology of Marek's Disease in Poultry
Marek's disease is highly contagious and readily transmitted among chickens. The virus matures into a fully infective, enveloped form in the epithelium of the feather follicle, from which it is released into the environment. It may survive for months in poultry house litter or dust. Dust or dander from infected chickens is particularly effective in transmission.
Once the virus is introduced into a chicken flock, regardless of vaccination status, infection spreads quickly from bird to bird. Infected chickens continue to be carriers for long periods and act as sources of infectious virus. Shedding of infectious virus can be decreased, but not prevented, by prior vaccination.
Severe clinical outbreaks of Marek's disease in commercial turkey flocks, with mortality rates from tumors reaching 40%–80% in chickens 8 to 17 weeks old, have been reported in France, Israel, and Germany.
Unlike virulent strains of Marek's disease virus, which are highly contagious, turkey herpesvirus is not readily transmissible among chickens (although it is easily transmitted among turkeys, its natural host).
Live, attenuated MDV strains vary greatly in their transmissibility among chickens; the most highly attenuated are not transmitted. Marek's disease virus is not vertically transmitted.
Clinical Findings of Marek's Disease in Poultry
The incidence of Marek's disease is quite variable in commercial flocks and depends on the following factors:
strain and dose of virus
age at exposure
maternal antibody
host sex and genetics
strain and dose of vaccine virus
several environmental factors, including stress
In addition to lymphoid neoplasms, Marek's disease virus can also induce other clinically distinct disease syndromes, including the following:
transient paralysis
early mortality syndrome
cytolytic infection
atherosclerosis
persistent neurologic disease
Typically, affected birds show only listlessness before death; however, a transient paralysis syndrome has been associated with acute MDV infection; chickens become ataxic for periods of several days and then recover. This syndrome is rare in immunized birds. Death is usually the result of paralysis, rendering the birds unable to reach food and water.
Lesions
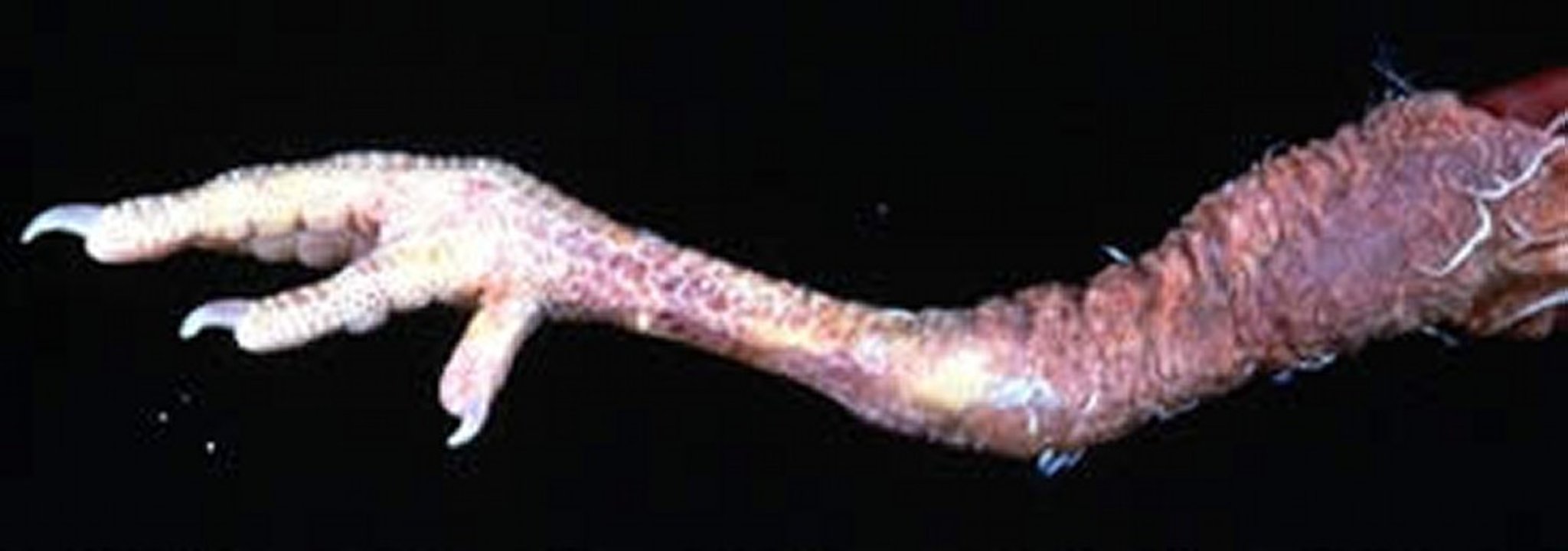
Enlarged nerves are one of the most consistent gross lesions in affected birds. Various peripheral nerves, but particularly the vagus, brachial, and sciatic, become enlarged and lose their striations (see and images).


Clinical signs associated with peripheral nerve dysfunction are frequently the most easily noticed. One leg stretched forward and one leg backward is a typical presentation of leg paresis in a chicken with Marek's disease.
Courtesy of Dr. Jean Sander.


Unilateral enlargement of sciatic nerve and sacral plexus in a chicken with Marek's disease.
Courtesy of Dr. John Dunn.
Diffuse or nodular lymphoid tumors may be observed in various organs, particularly the liver, spleen, gonads, heart, lung, kidney, muscle, and proventriculus. Enlarged feather follicles (commonly termed skin leukosis) may be noted in broilers after defeathering during processing and are a cause for condemnation. (See .)

Skin lesions are a common cause for condemnations of broiler chickens with Marek's disease. Broiler chickens can also have erythematous lesions of the shank, commonly referred to as Alabama red leg.
Courtesy of Dr. Jean Sander.
The cloacal bursa (bursa of Fabricius) is only rarely tumorous and more frequently is atrophic.
Histologically, the lesions consist of a mixed population of small, medium, and large lymphoid cells plus plasma cells and large anaplastic lymphoblasts. These cell populations undoubtedly include tumor cells and reactive inflammatory cells. When the cloacal bursa is involved, the tumor cells typically appear in interfollicular areas.
Diagnosis of Marek's Disease in Poultry
Standard criteria: history and clinical signs, gross pathological findings, and histological examination
Advanced criteria: immunohistochemical testing, standard and quantitative PCR assay, virus isolation, serological testing
For the diagnosis of Marek's disease, it is critical to diagnose the tumors and not the infection because MDV is considered ubiquitous within commercial poultry flocks. Usually, diagnosis is based on enlarged nerves and lymphoid tumors in various viscera.
The absence of bursal tumors helps distinguish this disease from lymphoid leukosis, although the presence of bursal tumors does not exclude Marek's disease.
Marek's disease can develop in chickens as young as 3 weeks old, whereas lymphoid leukosis typically is observed in chickens > 14 weeks old.
Reticuloendotheliosis, although rare, can easily be confused with Marek's disease, because both diseases can feature enlarged nerves and T-cell lymphomas in visceral organs.
Standard criteria are often sufficient for a presumptive diagnosis; however, advanced criteria are needed for a definitive diagnosis. Immunohistochemical testing can be used to confirm that tumors are composed of predominant T-cell populations or expressing specific MDV antigens. There is a quantitative association between viral load and Marek's disease tumors; most tumor-bearing chickens have high viremia titers and usually positive results of PCR assay. Thus, the demonstration of high quantities of virus, viral DNA, or viral antigens in tumor cells and the exclusion of other relevant tumor viruses should be sufficient for a specific diagnosis of Marek's disease.
Control of Marek's Disease in Poultry
Vaccination
Biosecurity
Breeding for genetic resistance
Vaccination is the central strategy for the prevention and control of Marek's disease, along with strict sanitation to decrease or delay exposure and breeding for genetic resistance.
The most widely used vaccines include the following:
Turkey herpesvirus (HVT, naturally avirulent Meleagrid alphaherpesvirus 1)
SB-1 or 301B/1 (naturally avirulent Gallid alphaherpesvirus 3)
CVI988/Rispens (attenuated Gallid alphaherpesvirus 2)
The HVT vaccine is now commonly used to create recombinant vaccines featuring the insertion of genes from other poultry viruses, such as Newcastle disease virus, infectious bursal disease virus, avian influenza H5 hemagglutinin, and infectious laryngotracheitis virus. These recombinant vaccines offer protection against both Marek's disease and the virus gene insert.
Bivalent vaccines consisting of HVT and either the SB-1 or 301B/1 strains of Gallid alphaherpesvirus 3 have been used to provide additional protection against challenge with virulent MDV isolates. Currently available commercial vaccines such as CVI988/Rispens and the new long terminal repeat (LTR) insertion derivatives appear to be the most protective against Marek's disease virus.
Vaccines are administered at hatch or in ovo to embryos at the 18th day of incubation. In ovo vaccination is now performed by automated technology and is widely used for vaccination of commercial broiler chickens, mainly because of decreased labor costs and greater precision of vaccine administration.
Proper handling of vaccine during thawing and reconstitution is crucial to ensure that adequate doses are administered.
Cell-associated vaccines are generally more effective than cell-free vaccines because they are less susceptible to inactivation by maternal antibodies. Under typical conditions, vaccine efficacy is usually > 90%.
Since the advent of vaccination, losses from Marek's disease have decreased dramatically in broiler and layer flocks. However, disease may become a serious problem in individual flocks or in selected geographic areas (eg, the Delmarva broiler industry). Of the many causes proposed for these excessive losses, early exposure to very virulent virus strains appears to be among the most important.
Key Points
Marek's disease is a highly contagious neoplastic disease characterized by T-cell lymphomas and peripheral nerve enlargement.
Leg paralysis is a common clinical sign
Presumptive diagnosis can be made based on history, clinical signs, gross pathological findings, and histological examination.
Vaccines are highly protective and should be used in combination with good biosecurity.
For More Information
Nair V, Gimeno I, Dunn J, et al. Neoplastic diseases. In: Swayne DE, ed. Boulianne M, Logue CM, McDougald LR, Nair V, Suarez DL, associate eds. Diseases of Poultry. 14th ed. Wiley; 2019:550-587.
Marek's disease. WOAH Terrestrial Manual. 2023.
Witter RL, Gimeno IM, Pandiri AR, Fadly AM. Tumor Diagnosis Manual: The Differential Diagnosis of Lymphoid and Myeloid Tumors in the Chicken. American Association of Avian Pathologists; 2010.





