



Nutritional management of the dairy cow has been intensively researched because milk and milk-based food products are an important part of human nutrition and meeting nutritional needs of the growing human population. The lactating dairy cow is an exceptional metabolic animal with very high nutritional requirements relative to most other species (see the table ). Meeting these requirements, especially for energy and protein, relative to intake capacity as controlled by dietary fiber, is challenging. Diets must have sufficient nutrient concentrations to support production and metabolic health while also maintaining rumen health and the efficiency of fermentative digestion.
The most recent publication describing dairy cattle nutrient requirements was released by the National Academies of Science, Engineering, and Medicine in December 2021, 20 years after the previous National Research Council publication (NRC 2001). This update (NASEM 2021) provides many nutrient requirement updates and new perspectives on feeding recommendations for dairy cows.
Feed Intake
The cornerstone of dairy nutrition is managing feed intake relative to absolute nutrient requirements. Feed intake, typically defined as dry matter intake (DMI), and feed efficiency (milk production [absolute or component corrected] per unit of DMI) are key nutritional monitoring metrics. DMI is influenced by the following factors:
Feed compositional factors: neutral detergent fiber (NDF) content, quality of ensiled feeds (excessive moisture and fermentation products), maturity (lignification) of forage, palatability attributes, rumen degradable protein, and nutrient availability
Cow physiological factors: age, body size, physiological state, body condition score, days in lactation, and production level
Management factors: feed bunk management (feed delivery, availability, and consistency), grouping strategies, cow comfort, water availability and quality, and heat abatement strategies
Lactating cows should be managed to maximize intake rapidly after calving to minimize the severity and duration of negative energy balance experienced. Milk production and associated energy requirements generally peak approximately 6–10 weeks into lactation, whereas DMI usually does not peak until 8–12 weeks into lactation. Severe postpartum negative energy balance will negatively impact body condition, resulting in greater risk for postpartum disease and reproductive inefficiency.
Intake regulation is a complex cerebral integration of signals from digestion products, gut distention, autonomic sensory inputs via the vagal nerve, and the milieu of metabolic regulatory hormones. Simplistically, intake is a function of the interaction between caloric intake and physical distention. Whether caloric status or physical distention has a greater impact on intake control depends upon the cow’s physiological state.
Feeding Guidelines for Large-Breed Dairy Cattle
Life Stagesa | Dry (Far-off) | Close (Close-up) | Cow - Fresh | Cow - Early | Cow - Mid | Cow - Late | 6-mo Heifer | 12-mo Heifer | 18-mo Heifer | 24-mo Heifer (Close-up) |
|---|---|---|---|---|---|---|---|---|---|---|
Body weight, kg | 740 | 740 | 700 | 675 | 725 | 740 | 200 | 300 | 450 | 595 |
DMI, kg/d | 13.9 | 12.3 | 20.8 | 25.8 | 29.4 | 27.4 | 5 | 7 | 9.8 | 11 |
Milk production, kg/db | — | — | 35 | 55 | 43 | 35 | — | — | — | — |
Crude protein, %c | 12 | 14.3 | 17.5 | 17.4 | 15.2 | 14.1 | 15 | 12.6 | 12 | 15.0 |
RDP, % | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 |
RUP, % | 1.9 | 4.3 | 7.5 | 7.4 | 5.2 | 4.1 | 5 | 2.6 | 2 | 5 |
MP, % | 5.2 | 6.7 | 10.9 | 10.2 | 10.2 | 10.1 | 8.1 | 7.0 | 6.1 | 14 |
NEL, Mcal/kg | 1.28 | 1.49 | 1.7d | 1.8 | 1.65 | 1.36 | — | — | — | |
ME, Mcal/kg | 2.26 | 2.09 | 1.92 | 2.12 | ||||||
NDF, min, % | 25–33 | 25–33 | 25–33 | 25–33 | 25–33 | 25–33 | 25–33 | 25–33 | 25–33 | 25–33 |
Forage NDF, min, % | 19–25 | 19–25 | 19–25 | 19–25 | 19–25 | 19–25 | 19–25 | 19–25 | 19–25 | 19–25 |
Starch, max, % | 15–20 | 15–20 | 22–30 | 22–30 | 22–30 | 22–30 | 15–20 | 15–20 | 15–20 | 15–20 |
Calcium, % | 0.31 | 0.39 | 0.64 | 0.64 | 0.60 | 0.58 | 0.78 | 0.58 | 0.44 | 0.39 |
Phosphorus, % | 0.19 | 0.21 | 0.39 | 0.39 | 0.37 | 0.35 | 0.32 | 0.23 | 0.21 | 0.19 |
Magnesium, % | 0.13 | 0.14 | 0.18 | 0.18 | 0.18 | 0.17 | 0.14 | 0.12 | 0.12 | 0.12 |
Chlorine, % | 0.13 | 0.14 | 0.34 | 0.34 | 0.32 | 0.29 | 0.17 | 0.14 | 0.13 | 0.13 |
Sodium, % | 0.16 | 0.7 | 0.23 | 0.22 | 0.22 | 0.21 | 0.17 | 0.15 | 0.15 | 0.16 |
Potassium, % | 0.62 | 0.69 | 1.10 | 1.07 | 1.00 | 0.99 | 0.51 | 0.54 | 0.56 | 0.60 |
Sulfur, % | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 |
Vitamin A, IU/kg | 5,850 | 6,630 | 3,687 | 3,021 | 3,303 | 3,103 | 3,390 | 4,265 | 4,698 | 5,288 |
Vitamin D, IU/kg | 1,442 | 1,595 | 1,085 | 1,085 | 952 | 1,021 | 924 | 1,044 | 1,281 | 1,442 |
Vitamin E, IU/kg | 85 | 181 | 22 | 22 | 19 | 20 | 49 | 56 | 68 | 77 |
Abbreviations: DMI, dry-matter intake; ME, metabolizable energy; MP, metabolizable protein; NDF, neutral detergent fiber; NEL, net energy lactation; RDP, rumen degradable protein; RUP, rumen undegraded protein. Trace mineral added to ration: cobalt, 0.2 mcg/g; copper, 8–19 mcg/g; iodine, 0.42–0.69 mcg/g; iron, 16–61 mcg/g; manganese, 27–49 mcg/g; selenium, 0.30 mcg/g; zinc, 30–66 mcg/g. | ||||||||||
Adapted, with permission, from National Academies of Science, Engineering, and Medicine (NASEM). Nutrient Requirements of Dairy Cattle. 8th rev ed. The National Academies Press; 2021. doi:10.17226/25806. | ||||||||||
aLife stages: far-off dry > 30 days from calving; close-up dry < 30 days from calving; fresh, early, mid, and late represent lactation periods. | ||||||||||
bMilk composition changes through lactation stages. | ||||||||||
cAll concentrations are on a dry-matter basis. | ||||||||||
dThe maximum energy concentration that can be formulated without resulting in insufficient dietary fiber or excess fat concentrations is 1.8 Mcal/kg dry matter. This energy concentration will typically not meet the energy requirements at this lactation stage, and these cows will lose weight. | ||||||||||
Dietary nonstructural carbohydrate content, through its fermentation to propionate, influences intake capacity in late pregnancy and immediately after calving. As the cow approaches peak milk production, physical distention, a function of dietary NDF content (see the table ) is the primary factor at peak milk production. Intake capacity of NDF varies with physiological state, ranging from 0.6% to 1.3% of body weight. Lowest intake capacity occurs in late pregnant heifers (0.6%) followed by late pregnant cows (0.8%). Physical fill becomes less of an intake-controlling factor in later lactation. Intake following peak milk production should be monitored to prevent excess body condition accumulation (body condition score > 3.5 on a 5-point scale).
Impact of Forage NDF on Forage Intake Capacitya
Forage Quality | Forage NDF, % | NDF Intake Capacity, % of body weight | ||||
|---|---|---|---|---|---|---|
0.6 | 0.8 | 1.0 | 1.2 | 1.4 | ||
Excellent | 38 | 1.58 | 2.11 | 2.63 | 3.16 | 3.68 |
42 | 1.43 | 1.90 | 2.38 | 2.86 | 3.33 | |
44 | 1.36 | 1.82 | 2.27 | 2.73 | 3.18 | |
46 | 1.30 | 1.74 | 2.17 | 2.61 | 3.04 | |
50 | 1.20 | 1.60 | 2.0 | 2.40 | 2.80 | |
54 | 1.11 | 1.48 | 1.85 | 2.22 | 2.59 | |
58 | 1.03 | 1.38 | 1.72 | 2.07 | 2.41 | |
62 | 0.97 | 1.29 | 1.61 | 1.94 | 2.26 | |
Poor | 66 | 0.91 | 1.21 | 1.52 | 1.82 | 2.12 |
Abbreviations: NDF, neutral detergent fiber. | ||||||
aValues shown are calculated forage intake as a percentage of body weight based on NDF intake capacity. | ||||||
Carbohydrates
Dietary carbohydrates comprise a wide range of compounds from simple sugars to complex starch and nonstarch polysaccharides. Carbohydrates account for 60–80% of dietary dry matter for dairy cows. Carbohydrate fractions are segregated based on chemical measures and nutritional impacts (see ). Complex polysaccharides associated with the plant cell wall that are more slowly fermented are quantified by measures of neutral detergent fiber (NDF), and its subfraction, acid detergent fiber (ADF). These structural carbohydrates limit intake but stimulate chewing and rumination, which helps maintain rumen buffering and health and can increase milk butterfat composition.

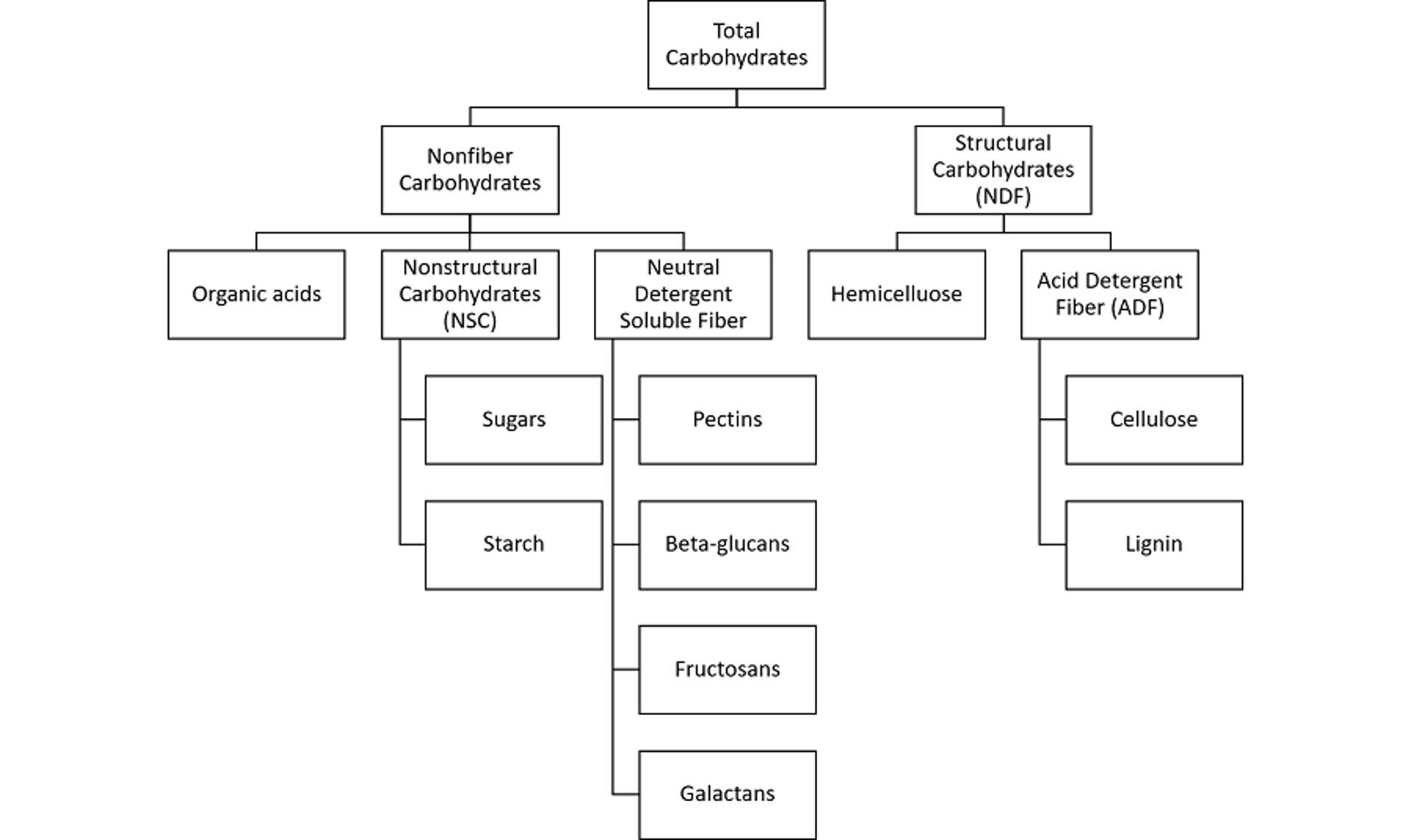
Diagram illustrating the organic composition of plant carbohydrates.
Courtesy of Dr. Robert Van Saun.
In general, fiber in the diet supports rumen health. Fiber in the rumen, especially fiber from forage sources that have not been finely chopped or ground, maintains rumen distention, which stimulates motility, cud chewing, and salivary flow. These actions affect the rumen environment favorably by stimulating the endogenous production of salivary buffers and a high rate of fluid movement through the rumen. Salivary buffers maintain rumen pH in a desirable range, while high fluid flow rates increase the efficiency of microbial energy and protein yield. Fiber, however, delivers less dietary energy than nonfiber carbohydrate (NFC). Fiber is generally less fermentable in the rumen than NFC, and rumen fermentation is the major mechanism by which energy is provided, both for the animal and the rumen microbes. Therefore, diets with high NDF concentrations promote rumen health but provide relatively less energy than diets high in NFC.
Feed NFC content is calculated by subtracting the proportions (as dry matter) of NDF, crude protein, fat, and ash from 100%. NFC primarily consists of organic acids, sugars, starch, and neutral detergent soluble fiber (NDSF). In fermented feeds, fermentation acids (ie, lactate, acetate, propionate) also contribute to the NFC fraction. The sum of sugars and starch is referred to as nonstructural carbohydrate (NSC), which should not be confused with NFC.
The NSC fraction is enzymatically digested and determined by measuring released glucose units. Fiber compounds associated with the secondary plant cell wall that are not digestible by mammalian enzymes but that are solubilized by neutral detergent are defined as NDSF. Although pectins, beta-glucans, and galactans are associated with the secondary plant cell wall, they are highly rumen fermentable and provide good sources of energy in the ruminant diet.
Balancing fiber and NFC/NSC fractions to optimize energy intake and rumen health is a challenging aspect of dairy nutrition. To increase the energy supply, dietary NDF concentrations are usually decreased by adding starch and other sources of NFC. This increases the rate and extent of rumen fermentation, which leads to greater energy availability. Increased ruminal fermentation also leads to the increased production of volatile fatty acids, which tends to lower rumen pH. At rumen pH < 6.2, fiber digestion is decreased; at pH ≤ 5.5, fiber digestion is severely diminished, feed intake may be decreased, and rumen health is generally compromised. Dietary starch content has a measurable impact on NDF fermentation, and the NASEM report adjusts potential dietary NDF fermentability relative to dietary starch content.
Recommended minimum NDF concentrations depend on the source and physical effectiveness of the NDF and the dietary concentration of NFCs. Fiber from forage sources is, in general, more effective at stimulating salivation and cud chewing than fiber from nonforage sources. Thus, one variable in the assessment of dietary NDF adequacy is the proportion of NDF contributed from forages. Minimum NDF concentrations in the diets for high-producing cows are 25–30%. When fiber sources from forage make up ≥ 75% of the NDF, then total NDF concentrations in the lower end of this range may be acceptable (see the table ). When a smaller portion of total NDF is derived from forage sources, total NDF concentrations should be in the upper end of this range.
Unfortunately, all dietary NDF is not equivalent relative to these biological actions. Historically, NDF sources have been characterized relative to their ability to stimulate milk fat production (eg, effective NDF [eNDF]) or stimulate chewing (eg, physically effective NDF [peNDF]). A new descriptor of dietary NDF, physically adjusted NDF (paNDF), has been developed and incorporated into the NASEM 2021 report.
The desired outcome of dietary paNDF is to maintain rumen pH at 6.0 or 6.1 to facilitate fiber fermentation. Determination of paNDF dietary content is based on interaction of dietary starch content, forage NDF (fNDF), and fiber source fragility determined by the ratio of ADF:NDF. Essentially, this modeling system is predicting the amount of dry NDF material to be retained on the 8-mm sieve of the Penn State Particle Separator based on inputs of dry NDF material retained on the 19-mm sieve, dietary starch, forage NDF contribution, and fiber fragility. As dietary starch increases, it will adversely affect NDF digestibility due to pH effects. Fiber fragility is used to estimate NDF retention within the rumen, further stimulating rumination. Low-fragility fiber from grasses will be retained in the rumen longer than more fragile fiber from legumes. Contribution of NDF from forage will also promote more rumination and potential buffering.
Maximum recommended NFC concentrations are 38–44%. Diets with higher NFC concentrations will benefit from higher proportions of NDF coming from forage sources. These recommendations must be viewed as broad guidelines rather than strict rules. Factors including the total fermentability of the diet and the fermentability of the NDF influence the NDF requirement. Diets with highly fermentable NDF sources require higher total concentrations of NDF; however, they provide more energy per mass unit of NDF than diets with less fermentable NDF. Feeding management schemes such as total mixed rations result in lower minimum NDF concentrations than feeding dietary components individually.
Recommended Minimum NDF Concentrations Based on Proportion of NDF Coming From Forage Sources
NDF From Forage (% of Dietary Dry Matter) | NDF From Forage (% of NDF) | Minimum Total NDF (% of Dietary Dry Matter) | Maximum Dietary Starch (% of Dietary Dry Matter) |
|---|---|---|---|
19 | 75 | 25 | 30 |
18 | 66 | 27 | 28 |
17 | 58 | 29 | 26 |
16 | 52 | 31 | 24 |
15 | 45 | 33 | 22 |
Abbreviations: NDF, neutral detergent fiber. As the proportion of NDF from forage sources decreases, the minimum requirement for total NDF increases. These represent minimum requirements; diets with higher NDF concentrations present no problem and are generally appropriate for animals with relatively low energy requirements. Adapted, with permission, from National Academies of Science, Engineering, and Medicine (NASEM). Nutrient Requirements of Dairy Cattle. 8th rev ed. The National Academies Press; 2021. doi:10.17226/25806 | |||
Energy
Dietary energy available for metabolic use is referred to as metabolizable energy (ME). In contrast to digestible energy (DE), the feed energy losses in urine and fermentation gases are subtracted. The efficiency of ME utilization varies based on the physiological functions supported, which include body maintenance, growth, and lactation.
The net energy (NE) system takes into account the differences in efficiency of ME utilization for each of these processes and assigns a separate NE value to individual feedstuffs based on each of these energy-requiring processes (ie, body maintenance, growth, and lactation). Thus, in the US, in which the NE system is typically used, energy values of feedstuffs for ruminants are expressed as NE for maintenance (NEM), NE for gain (NEG), and NE for lactation (NEL). This system is cumbersome and nonintuitive and has many computational disadvantages compared with alternative systems based directly on ME. However, the NE system has the major advantage of more equitably comparing the energy values of forages to concentrates when used in ruminant diets.
The table has typical values for ME, NEL, NEM, and NEG for some feedstuffs commonly fed to dairy cows.
Dry Matter, Energy, Crude Protein, Fiber, and Nonfiber Carbohydrate Concentrations of Some Feedstuffs Commonly Fed to Dairy Cattlea
Dry Matter, % | Metabolizable Energy, Mcal/kg | Net Energy for Lactation, Mcal/kg | Net Energy for Maintenance, Mcal/kg | Net Energy for Gain, Mcal/kg | Neutral Detergent Fiber, % | Nonfiber Carbohydrate, % | Crude Protein, % | |
|---|---|---|---|---|---|---|---|---|
Ground shelled corn | 88 | 3.12 | 2.01 | 2.16 | 1.48 | 9.5 | 75.47 | 9.4 |
Corn silage | 35 | 2.33 | 1.45 | 1.57 | 0.97 | 45 | 40 | 8.8 |
Pasture | 20 | 2.46 | 1.54 | 1.67 | 1.06 | 45.8 | 19.6 | 26 |
Hay (cool-season grasses) | 84 | 2.02 | 1.23 | 1.33 | 0.75 | 58 | 21.6 | 13 |
Hay crop silage (cool-season grasses) | 42 | 1.92 | 1.16 | 1.25 | 0.68 | 58 | 18.2 | 17 |
Grass-legume mix hay | 85 | 2.07 | 1.25 | 1.35 | 0.77 | 50.8 | 23.8 | 18.4 |
Grass-legume mix hay crop silage | 44 | 1.96 | 1.23 | 1.32 | 0.74 | 50.4 | 21.5 | 19 |
Legume hay | 84 | 2.09 | 1.28 | 1.38 | 0.8 | 43 | 27.3 | 20.8 |
Legume hay crop silage | 43 | 2.01 | 1.22 | 1.32 | 0.74 | 43 | 25.1 | 22 |
Oats (rolled) | 90 | 2.78 | 1.77 | 1.9 | 1.26 | 30 | 50.2 | 13.2 |
Sorghum silage | 29 | 1.85 | 1.11 | 1.18 | 0.62 | 61 | 21.9 | 9.1 |
Soybean meal (solvent extracted, 44% crude protein) | 89 | 3.31 | 2.13 | 2.29 | 1.59 | 15 | 27.6 | 49.9 |
Cottonseeds | 90 | 2.91 | 1.94 | 1.96 | 1.31 | 50.3 | 5.1 | 23.5 |
Dried distillers' grains with solubles | 90 | 3.03 | 1.97 | 2.07 | 1.41 | 39 | 24.4 | 30 |
Corn and cob meal | 89.2 | 2.91 | 1.86 | 2 | 1.35 | 21.5 | 65.2 | 8.6 |
Adapted, with permission, from National Academies of Science, Engineering, and Medicine (NASEM). Nutrient Requirements of Dairy Cattle. 8th rev ed. The National Academies Press; 2021. doi:10.17226/25806 aEnergy values are estimated based on a consumption rate 3 times the rate of consumption at maintenance. These are representative values intended primarily for relative comparison among feeds. Analyzed values of actual feed samples will vary substantially, especially among forages. Laboratory analyses of feeds and forages are always advisable for both comparative evaluation and ration balancing. | ||||||||
Energy values for feeds are not directly measured by feed analysis laboratories but rather predicted using regression equations. Direct measurement of feed energy is only achieved using calorimetry methods; however, feeding a single feed is not practical. Feed energy values reported on laboratory feed analysis reports are estimates, usually based on formulas with ADF concentration and possibly crude protein as primary independent variables.
In the US, energy requirements of adult dairy cows are typically expressed in terms of NEL. This applies to pregnant dry cows as well as lactating animals. Maintenance requirements for mature cows of various mature body weights are given in the table . Energy requirements per kilogram of milk produced at various milk fat concentrations are given in the table .
Maintenance Energy Requirements for Cows of Various Body Weights
Body Weight, kg | Daily Energy Requirement, Mcal NEL/d |
|---|---|
400 | 8.94 |
450 | 9.77 |
500 | 10.57 |
550 | 11.36 |
600 | 12.12 |
650 | 12.87 |
700 | 13.61 |
750 | 14.33 |
800 | 15.04 |
Abbreviations: NE, net energy. |
Dietary Net Energy Requirement for Milk Production
Milk Fat Concentration, % | Milk True Protein Concentration, % | Dietary Energy Requirement/kg Milk Produced, Mcal NELa | |
|---|---|---|---|
Based on Milk Fat % | Based on Milk Fat and True Protein | ||
3.5 | 2.69 | 0.70 | 0.67 |
4 | 3.08 | 0.75 | 0.74 |
4.5 | 3.46 | 0.80 | 0.81 |
5 | 3.85 | 0.84 | 0.88 |
5.5 | 4.23 | 0.89 | 0.95 |
6 | 4.62 | 0.94 | 1.02 |
6.5 | 4.85 | 0.99 | 1.09 |
Abbreviations: NE, net energy. aEquations predicting milk energy content from either milk fat percentage alone or milk fat and milk true protein, assuming a set milk lactose content of 4.85% and milk fat:true protein ratio of 1.3:1. | |||
The required dietary energy concentration is a function of the energy requirement and feed intake rate. Calculated requirements for dietary energy concentration typically are very high in early lactation because the rate of milk production is high relative to the feed intake rate. However, the ration energy density concentrations required to meet the energy requirement of cows in very early lactation may be too high to be compatible with adequate dietary fiber concentrations. In general, diets with energy concentrations > 1.71–1.76 Mcal/kg do not contain adequate fiber to support good rumen health and function. Thus, dairy cows in early lactation typically cannot meet their energy requirements and are expected to lose weight.
Pearls & Pitfalls
|
Fats
Ruminant diets typically are low in total fat content (< 5% dry matter) due to the negative effects that polyunsaturated fatty acids (PUFAs) have on microbial fiber fermentation. Dietary fat is derived from three sources:
Endogenous fats: forage lipids that include glycolipids, pigments, cutins, and waxes
Vegetable fats: polyunsaturated fats from oilseeds such as soybean, corn, canola, sunflower, and flaxseed
Rumen inert fats: saturated animal fats, calcium soaps, and prilled fats
Typically, each type of fat source can be supplied in the diet at 2–3% of dry matter up to a total of 8–9% total fat. Fats in ruminant diets can induce undesirable metabolic effects, both within the rumen microbial population and within the animal. Ramifications of these effects include decreased fiber digestion, indigestion with poor rumen health, suppression of milk fat concentration (ie, milk fat depression), or some combination.
Quantifying rumen unsaturated fatty acid load (RUFAL) is another method of evaluating dietary fat and potential impacts on rumen microbes or cow metabolism and fat production. Dietary PUFAs are naturally biohydrogenated by rumen microbes to generate saturated fatty acids, which accounts for ruminant body fat being predominately saturated. Dietary situations in which higher dietary PUFAs are being fed, coupled with lower rumen pH, result in an alternative biohydrogenation pathway in the rumen leading to bioactive conjugated linoleic acid (CLA) production. One of these CLA isomers (trans-10, cis-12 CLA) inhibits mammary de novo fat synthesis, resulting in milk fat depression (milk fat < 3%).
Supplemental dietary fat can provide additional concentrated energy to meet the lactating cow's energy needs. Adding fat within the first 3 weeks of lactation potentially can decrease feed intake, which is not desirable in early lactation. The addition of fat after this period may improve milk fat content, milk production, or reproductive efficiency; however, response to dietary fat supplementation is not consistently predictable. Historically, the amount of total dietary fat consumed was suggested to be limited to the amount of fat produced (milk production × fat percentage); however, the contribution from dietary PUFA is more important and should be limited.
Protein
Lactating dairy cow protein requirements are based on amino acids required for maintenance and milk protein synthesis, with additional amino acids to support growth in the first two lactations. Ruminant animals derive from microbial protein most of the amino acids that support body metabolism. Microbial protein is of high biological value and highly digestible. Mixed microbes contain between 45% and 60% crude protein. Therefore, dietary formulations are directed to ensure optimum microbial growth to minimize the need for expensive supplemental dietary protein.
Dietary protein not degraded by rumen microbes can be potentially digested in the abomasum and resultant amino acids absorbed in the small intestine. This fraction of dietary protein is termed rumen-undegradable protein (RUP), in contrast to the dietary protein fraction degraded in the rumen (rumen-degradable protein [RDP]) and used by the microbes. To their advantage, ruminants are able to use low-quality protein or nonprotein nitrogen (NPN) sources to support microbial protein synthesis in meeting their amino acid requirements. Ruminants consuming low-protein diets will efficiently recycle blood urea via saliva to the rumen to provide needed nitrogen sources to support microbial populations.
Two systems of describing the dietary protein supply and requirements for dairy cows are in general use:
Crude protein system: based on dietary nitrogen converted to protein equivalent using 6.25 multiplier factor (assumes protein is 16% nitrogen); does not account for differences in availability to rumen or cow
Metabolizable protein system: based on a dynamic rumen model describing potential dietary nitrogen and carbohydrate pools available to rumen microflora. Based on dietary parameters and defined rates of passage and digestion interactions, microbial protein flow and dietary RUP fraction are predicted. Digestible amounts of microbial protein and RUP are predicted and collectively account for total metabolizable protein (MP) delivered to the small intestine and absorbed.
The crude protein system is relatively simple to use and historically has been the traditional means of formulating dairy cow rations. The table provides general guidelines for the required crude protein concentration of diets for large- and small-breed dairy cattle at various levels of production. It can be used for general evaluations of the protein adequacy of dairy diets.
The MP system is more complex than the crude protein system. It was developed in recognition of the need to provide dietary nitrogen to support microbial growth (ie, RDP) in addition to dietary RUP to collectively meet cow amino acid needs. Unlike crude protein, MP cannot be directly measured in a feed ingredient via laboratory analysis and must be estimated using dynamic rumen models.
See also the graphic depicting the .
Estimated Minimum Dietary Protein Concentrations for Dairy Cows at Various Levels of Production
Dietary Protein Requirements, % of Dry Matter | ||||
|---|---|---|---|---|
__________________________________________________________________________________________ | ||||
Large-Breed Cows | Small-Breed Cows | |||
______________________________________________ | _______________________________________________ | |||
Milk production, kg/d | Crude Proteina | Metabolizable Proteinb | Crude Proteina | Metabolizable Proteinb |
18 | — | — | 15.0 | 12.9 |
23 | — | — | 16.4 | 13.1 |
27 | 14.5 | 11.0 | 17.5 | 13.3 |
32 | 15.0 | 11.2 | 18.4 | 13.3 |
36 | 15.8 | 11.5 | 19.0 | 13.3 |
40 | 16.5 | 11.7 | — | — |
45 | 17.3 | 11.9 | — | — |
50 | 17.8 | 12.0 | — | — |
55 | 18.3 | 12.1 | — | — |
This table is intended to provide a general reference for initial evaluations of dairy diets, not a standard against which rations are balanced. Calculation of dietary metabolizable protein concentrations generally requires specialized software. | ||||
aCrude protein requirements were generated by Spartan Dairy version 3.0 (Michigan State University), assuming a mature cow in mid lactation. | ||||
bMetabolizable protein (MP) requirements were generated by NRC Model Software for Nutrient Requirements of Dairy Cattle (National Research Council, 2001). Actual MP will depend upon dietary composition and stage of lactation as milk composition changes. | ||||
The efficiency with which RDP is recovered as microbial protein depends on the growth rate of the rumen microbes, which in turn depends on the supply of fermentable energy sources in the rumen. Thus, diets with sufficient RDP and relatively high energy concentrations will result in high yields of microbial protein, which will become available for intestinal digestion and absorption as MP. Calculations that balance dairy diets for MP must consider the complex interrelations among fermentable energy sources, RDP, and RUP. In general, specialized software, commercially available, is necessary to formulate dairy diets using the MP system. Even with such software, many variables must be estimated with uncertainty. Therefore, calculations of MP supply must be recognized to be approximations.
Relationship of dietary protein intake to metabolizable protein supply
Diagram showing the relationship of dietary protein intake to metabolizable protein supply. The two branch points (indicated by 1 and 2) constitute the major variables relating the dietary crude protein supply to the metabolizable protein supply. The first branch point represents the proportion of protein that is degraded in the rumen. This branch point is influenced by inherent properties of the protein and the rate of ingesta passage through the rumen. The second branch point represents the proportion of nitrogen from degraded protein that is recaptured as microbial protein. This is influenced by the microbial growth rate, which depends on the supply of rumen available energy. Nitrogen that is not recaptured as microbial protein is absorbed from the rumen as ammonia and converted to urea by the liver. Some urea is recycled back to the rumen; however, a large portion is excreted in urine. MCP, metabolizable crude protein; MP, metabolizable protein; N, nitrogen; RDP, rumen degraded protein; RUP, rumen undegraded protein. |

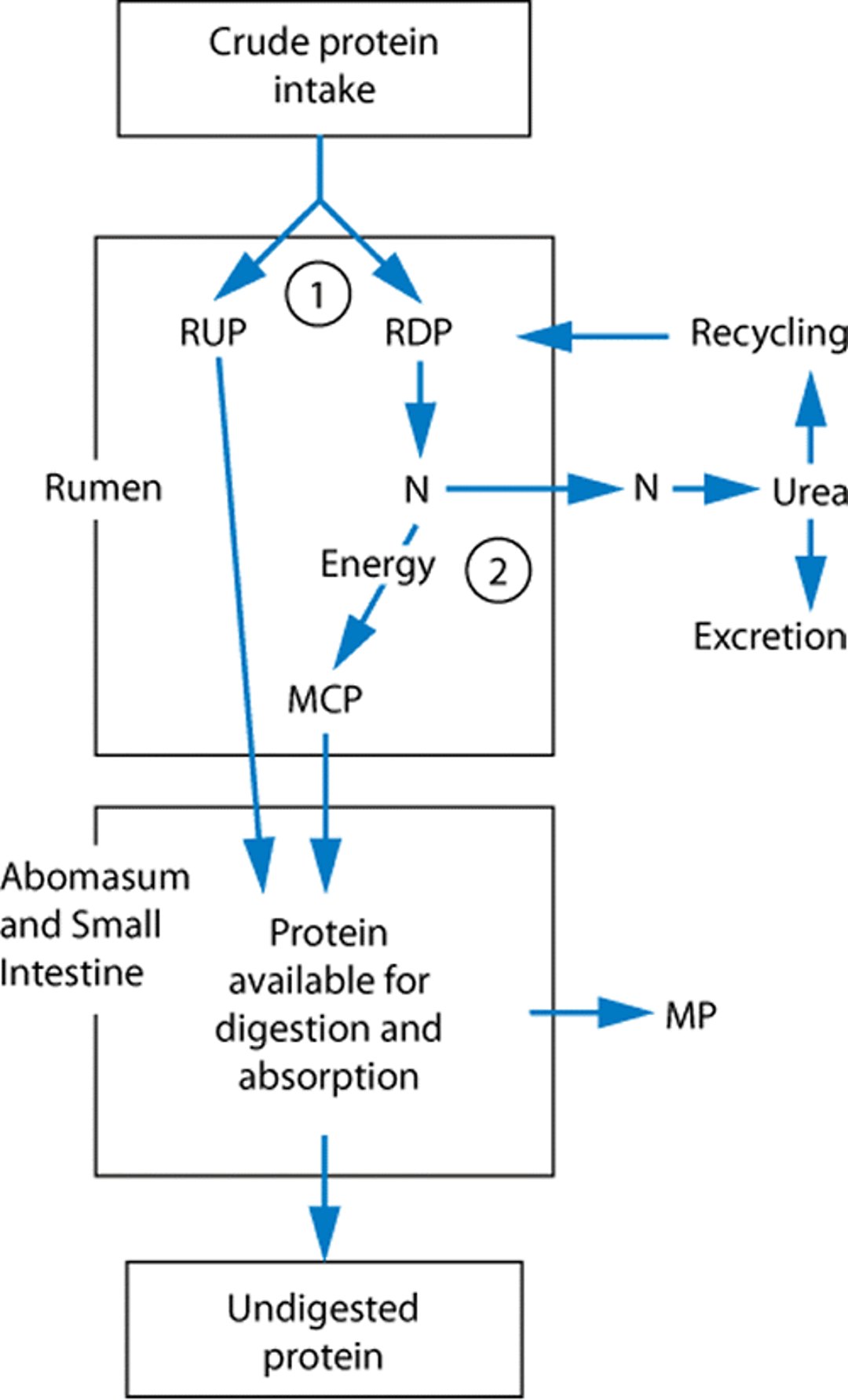
Dietary ingredients vary in their proportion of RDP and RUP. In general, feeds with high moisture and high protein concentrations (eg, legume silages) have a high proportion of RDP. In contrast, feeds that have been processed, and especially those that have undergone drying or roasting, have relatively higher proportions of RUP. The percentage of RDP and RUP for a feed will always equal 100%; however, the proportion of each for any ingredient is not fixed and will be altered by changes in rates of passage through the rumen. Intake rate, body size, and dietary fiber amount and effectiveness will influence rate of passage.
At high rates of feed intake, the rate of feed passage through the rumen is high; thus, there is less opportunity for rumen protein degradation than with the same feeds at lower intake rates. Therefore, with cows on the same diet, RUP proportions are higher in animals with high rates of feed intake than in those with low rates of feed intake. Animals most likely to benefit from supplements selected for high RUP proportions are those with relatively high protein requirements and relatively low rates of feed intake.
The specific amino acid requirements of dairy cows are not as well understood as those of swine or poultry. Most research has focused on methionine and lysine as first limiting amino acids in typical dairy cattle diets, especially during early lactation. High-forage diets in which the cow relies primarily on microbial protein may be deficient in histidine. Insufficient amino acids can be supplemented by feeding targeted RUP ingredients high in these amino acids or rumen-protected forms of these amino acids. The NASEM 2021 software estimates amino acid supply and efficiency of utilization for dairy cows on different diets. With typical feedstuffs, if the MP requirement is met and the dietary lysine:methionine ratio is approximately 3:1, then the amino acid requirements for milk production are likely being optimized.
Water
The availability of high-quality water for ad libitum consumption is critical. Insufficient water intake leads immediately to decreased feed intake and milk production. Water requirements of dairy cows are related to milk production, DMI, ration dry-matter concentration, salt or sodium intake, and ambient temperature.
Various formulas have been devised to predict water requirements. For recommended formulas to estimate water consumption of lactating and dry dairy cows, see the table .
Free Water Intake Equations for Dairy Cows
Cow Status | Equation |
|---|---|
Lactating cows | |
— Equation 1 | FWI (L/d) = −91.1 + (2.93 × DMI) + (0.61 × DM%) + (0.062 × NaK) + (2.49 × CP%) + (0.76 × Temp) |
— Equation 2 | FWI (L/d) = −60.2 + (1.58 × Milk) + (0.064 × NaK) + (0.83 × DM%) + (0.54 × Temp) + (0.08 × DIM) |
Dry cows | |
FWI (L/d) = (1.16 × DMI) + (0.23 × DM%) + (0.44 × Temp) + (0.061 × [Temp − 16.4]2) | |
Abbreviations: CP%, dietary crude protein (% dry matter); DIM, days in milk; DM%, dietary dry matter content; DMI, dry matter intake (kg/d); FWI, free water intake; Milk, daily milk yield, kg/d; NaK, sum of dietary Na plus K (mEq/kg dry matter); Temp, dairy mean ambient temperature (°C). | |
Adapted, with permission, from National Academies of Science, Engineering, and Medicine (NASEM). Nutrient Requirements of Dairy Cattle. 8th rev ed. The National Academies Press; 2021. doi:10.17226/25806 | |
Providing adequate access to water is critical to encourage maximal water intake. Water should be placed near feed sources and in milking parlor return alleys because most water is consumed in association with feeding or after milking. For water troughs, a minimum of 5 cm (2 inches) of length per cow at a height of 90 cm (3 feet) is recommended. One water cup per 10 cows is recommended when cows are housed in groups and given water via drinking cups or fountains. Every cow group should have a minimum of two watering stations to prevent a high social order cow from blocking a single water source.
Individual cow water intake rates are 4–15 L/min, anytime a cow drinks during a day. Many cows may drink simultaneously, especially right after milking, so trough volumes and drinking cup flow rates should be sufficient to ensure water availability during times of peak demand. Water troughs and drinking cups should be cleaned frequently and positioned to avoid fecal contamination.
Poor water quality may result in decreased water consumption, with resultant decreases in feed consumption and milk production. Water can be evaluated by its organoleptic properties (color, taste, and smell) or quantification of dissolved or suspended contents. Factors affecting water quality include the following:
pH: A wide pH range from 5 to 9 seems acceptable to cattle; extremes of the pH range may be of concern for palatability.
Microbiological contamination: Bacterial counts that may cause digestive issues in ruminants have not been well documented. No correlation has been found between bacterial contamination level and cow performance; however, if bacteria are present, cleaning watering units more frequently is reasonable.
Total dissolved solids (TDS): Also referred to as total soluble salts, TDS is a major factor that refers to the total amount of inorganic solute in the water. TDS is generally expressed as mg/L or parts per million (ppm), which are numerically equivalent values (see the table ).
Hardness: A measure of calcium and magnesium content in water; not equivalent to TDS; generally not shown to affect cow performance; however, calcium may add to the amount in the diet.
Mineral content: Water can contain a range of mineral elements that are both essential nutrients and toxic elements. The table lists potential elemental contaminants of drinking water with upper-limit guidelines.
Inorganic contaminants: Beyond mineral elements, other inorganic contaminants of concern for ruminants include nitrates, nitrites, and sulfates. Concentrations of nitrate (expressed as nitrate nitrogen) < 10 mg/L are safe for ruminants. At concentrations > 20 mg/L, cattle may be at risk, especially if nitrate concentrations in the feed are high. Water with nitrate concentrations > 40 mg/L should be avoided. General recommendations for sulfate concentrations in drinking water are < 500 mg/L for calves and < 1,000 mg/L for adult cattle. The specific sulfate salts present in water may affect the response of cattle; iron sulfate is the most potent depressor of water intake.
Organic contaminants: A wide range of organic compounds such as herbicides, insecticides, and pharmacological agents may contaminate water sources. Industrial pollution from mining and gas drilling may increase solvents, fuels, methane, and other surfactants or chemicals.
Water testing is readily available; however, to evaluate water for food-producing animals, identifying an appropriate laboratory that addresses this specifically is best. Contact the laboratory for directions on appropriate sampling technique and sample containers.
Guidelines for Total Soluble Salts (Total Dissolved Solids) in Drinking Water for Cattle
Total Soluble Salts, mg/L | Comments |
|---|---|
< 1,000 | Safe and should pose no health problems. |
1,000–2,999 | Generally safe but may cause a mild temporary diarrhea in animals not accustomed to the water. |
3,000–4,999 | Water may be refused when first offered to animals or cause temporary diarrhea. Animal performance may be less than optimum because water intake is not maximized. |
5,000–6,999 | Pregnant or lactating animals should not drink such water. May be offered with reasonable safety to animals when maximum performance is not required. |
≥ 7,000 | These waters should not be offered to cattle. Health problems and/or poor production will result. |
Adapted, with permission, from National Research Council. Nutrient Requirements of Dairy Cattle. 8th rev ed. The National Academies Press; 2001. doi:10.17226/9825 | |
The table lists potential elemental contaminants of drinking water with upper-limit guidelines.
Safe Drinking Water Guidelines for Cattle
Element | Potentially Concernable Concentrations for Cattle, mg/L |
|---|---|
Aluminum | 5–10 |
Arsenic | 0.20 |
Boron | 150 |
Cadmium | 0.01–0.05 |
Chromium | 0.1–1.0 |
Cobalt | 1.0 |
Copper | 1.3 |
Fluorine | 2.0 |
Lead | 0.05–0.10 |
Manganese | 0.50 |
Mercury | 0.01 |
Nickel | 1.0 |
Selenium | 0.05 |
Vanadium | 0.1 |
Zinc | 5.0–25 |
Adapted, with permission, from National Academies of Science, Engineering, and Medicine (NASEM). Nutrient Requirements of Dairy Cattle. 8th rev ed. The National Academies Press; 2021; doi:10.17226/25806 | |
Minerals
Calcium and Phosphorus Requirements of Dairy Cattle
Of the macrominerals required by dairy cattle, calcium and phosphorus are most often considered due to their roles in skeletal structure, metabolism, and milk production.
Calcium requirements of lactating dairy cows are high relative to other species and to nonlactating cows because of the high calcium concentration (1.1–1.25 g/kg) in milk.
Phosphorus requirements are approximately half of calcium requirements; phosphorus is lost to milk (0.8–1.0 g/kg) as well as recycled via saliva to the rumen to support microbial growth needs. Phosphorus is considered the first limiting mineral on a forage-based feeding program due to the low phosphorus content of forages (< 0.21% dry matter), especially without fertilization. Phosphorus has been associated with infertility in cattle; however, this occurs only in extremely low forage phosphorus (< 0.15% dry matter) conditions. Of greater concern is phosphorus oversupplementation and its environmental impact.
Due to its tight homeostatic control, blood calcium concentration is very consistent within cows of specific age groups. Blood phosphorus concentration in preweaned calves is higher than in adult cows, with a decrease in concentration when phosphorus starts to be recycled through the saliva to the rumen. Consequently, blood calcium:phosphorus ratio is approximately 1.2–1.5:1 in younger animals, compared to 1.5–2:1 in older animals. Dietary calcium and phosphorus supplementation amounts and ratio are important in young growing animals in support of bone development and in lactating cows for milk production. Early lactation often results in a period of negative calcium and phosphorus balance resulting in bone mobilization.
Legume and grass forages have similar phosphorus content but divergent calcium content, with legumes having much higher calcium levels. Depending on the amount of legume forage in the diet, calcium may need to be supplemented to support lactation. More recent requirement models account for differences in bioavailability of a mineral element depending on the ingredient source. Inorganic mineral sources have the highest availability (75–90%), though there is some variability among different mineral sources. Forages have a lower availability for calcium (30%) and phosphorus (64%), resulting from chelating compounds such as phytic acid and oxalic acid found in plant tissues. Mineral requirements are presented as total and available, with the difference being the mineral availability from various dietary ingredients being considered.
Beyond their support for growth and lactation, nutrition of calcium and phosphorus is of great interest relative to clinical (ie, milk fever) and subclinical hypocalcemia. The role of each mineral has been researched extensively in determining methods of preventing parturient paresis. Initially, dietary calcium:phosphorus ratio was considered, though this usually resulted in supplementing more dietary phosphorus to balance against the high endogenous dietary calcium. Research indicates that restricting dietary calcium at or below the available requirement 2–3 weeks before calving induces the homeostatic system to upregulate calcium influx to counter subsequent colostrum and milk calcium losses. This approach requires diets with calcium concentrations near 0.3% of dry matter. Such diets are difficult to formulate with available feedstuffs while still meeting other nutritional requirements.
Another preventive approach to hypocalcemia utilizes the effect of acid-base balance on mobilizing body calcium. This approach uses dietary cation-anion difference (DCAD) as a method of inducing renal compensated metabolic acidosis. Research has shown that renal activation of vitamin D via parathyroid hormone stimulation is facilitated when there is a state of slightly lower blood pH compared to a more alkaline state. Multiple equations determining DCAD have been proposed, based on strong ion contributions to acid-base balance. All equations have been validated relative to manipulating DCAD and hypocalcemia prevalence. The following are the two most commonly used equations:
(1) DCAD, mEq/kg dry matter = (Na + K) – (Cl + S) and
(2) DCAD, mEq/kg dry matter = (Na + K) – (Cl + 0.6 × S),
where the dietary content (% dry matter) of these minerals needs to be converted to mEq/kg dry matter. This is accomplished by multiplying Na, K, Cl, and S dietary content (% of dry matter) by 434.8, 256.4, 281.7, and 625, respectively. Research and field experience have shown that this dietary approach is effective for hypocalcemia prevention; however, adoption on farms is variable due to the need to monitor response, economics, and logistics for incorporating this method into the dry cow diet.
Dietary calcium content with the DCAD approach to milk fever prevention has been debated. Recent research has suggested that higher dietary calcium is not necessary to compensate for increased urinary calcium loss (1, 2). However, other studies also show high calcium diets can be beneficial with DCAD (3,4). Collectively, the research would indicate dietary calcium between 0.5% and 1.2% (dry-matter basis) is sufficient.
A new approach to hypocalcemia prevention addressing dietary calcium content has been proposed. A feed additive initially believed to bind dietary calcium, potentially decreasing calcium availability and stimulating the homeostatic system, has been studied. These studies found that the feed additive was actually binding phosphorus, and, to a lesser extent, magnesium (5,6,7). These findings were consistent with other research suggesting lower dietary phosphorus could improve calcium homeostasis at calving. Recommended diet phosphorus concentration in dry cow diets ranges from 0.25% to 0.35% (dry-matter basis). High dietary phosphorus (> 0.5%) may promote hypocalcemia and contribute to environmental pollution.
Serum concentrations of calcium and inorganic phosphorus are of value in assessing the short-term homeostasis of these minerals; however, they are of little value in assessing long-term nutritional status. Bone ash concentrations are the best way to assess long-term calcium and phosphorus nutritional status.
Other Macromineral Requirements of Dairy Cattle
Other macrominerals of consequence to dairy cows include sodium, chloride, potassium, magnesium, and sulfur.
Sulfur is required primarily to provide substrate to rumen microbes in generating sulfur-containing amino acids as part of microbial protein generation. The ratio of nitrogen to sulfur in microbial protein is 14.5:1. A typical recommendation for ruminant diets is to maintain nitrogen:sulfur ratio between 10:1 and 12:1. A more biological approach is to estimate sulfur needed for microbial protein synthesis based on dietary metabolizable energy and protein availability. Sulfur generally is not deficient in the diet unless forages are grown on sulfur-deficient soils. Recommended dietary concentration is 0.21–0.25% dry matter.
The other macrominerals serve important biological roles as electrolytes located either extracellularly (sodium and chloride) or intracellularly (potassium and magnesium). These electrolytes have roles in acid-base balance, cell membrane electrical potentials, nerve conduction, and active transport.
Sodium needs to be supplemented in the diet as sodium chloride (eg, NaCl, common salt) or provided as free-choice salt. Forage contains very little sodium (< 0.05% dry matter) unless contaminated with salty water. Cows have an appetite for salt and will seek it out; thus, salt is used as a carrier for trace mineral supplements. Signs of severe salt deficiency include licking and chewing on fences and other environmental objects, urine drinking, and general ill thrift.
Insufficient dietary sodium results in decreased feed intake with subsequent decreases in animal performance. Milk production is decreased within 1–2 weeks of removing supplemental salt from the diets of lactating cows. Completely withholding salt from dry cow diets to prevent udder edema at calving is not a good feeding practice. Additional salt is necessary during heat stress. Sodium requirement ranges from 0.15% (dry cow) to 0.23% (lactating cow) dry matter.
Chloride typically follows sodium in biological systems, commonly added to diets as NaCl; additional chloride without sodium is used in diets to supplement more anions to decrease DCAD for dry cow diets.
Chloride requirement ranges from 0.25% to 0.29% dry matter.
Potassium plays an important role as the primary intracellular cation. Supplementing potassium in the diet is typically not necessary given the high potassium content of forages (grasses and legumes), especially with excess potassium (ie, potash) soil fertilization. Excess dietary potassium plays an important role in acid-base status that interferes with calcium homeostasis. Additional dietary potassium may be appropriate in early lactation and during heat stress to counter metabolic acid load. Potassium requirement ranges from 0.6% to 1.2% dry matter.
Magnesium may need to be fed with diets containing high proportions of grass forages, especially those consisting of rapidly growing pasture grasses. Such forages typically have low magnesium concentrations as well as high concentrations of potassium and organic acids, which interfere with the availability of dietary magnesium. Magnesium oxide is the typical magnesium supplement in ruminant diets. Magnesium requirement is between 0.21% and 0.35% dry matter, depending on dietary potassium.
Trace Mineral Requirements of Dairy Cattle
The trace minerals typically supplemented or measured in dairy cow diets include cobalt, copper, iron, manganese, selenium, iodine, and zinc. Trace minerals are required in minute amounts on the order of micrograms or nanograms per day. These minerals play important biological roles as components of metalloenzymes influencing metabolic reactions, immune function, and antioxidant status (see the table ).
Ruminants are challenged relative to trace mineral nutrition. Although there is great geographic variation in forage trace mineral status, most are marginal to low, generally resulting in a deficient diet without supplementation. Additionally, rumen microbial environment can alter these metal cations to decrease their valence and thus absorption efficiency at the small intestine. Rumen microbes generate chelating ligands to also prevent absorption. In the 2001 NRC report, bioavailability estimates (ie, absorption coefficients) for ingredient sources were provided. The 2021 NASEM report has further refined trace mineral source absorption coefficients based on newer data. One important issue with minerals that has not been incorporated into NASEM 2021 is the documented mineral antagonistic interactions; thus, there remains a need to be aware of these in any given diet and accounted for in rate of supplementation.
Micromineral Functions, Deficiency Diseases, and Interrelationships
Micromineral | Requirement, mg/kg dry matter | Major Biological Function | Deficiency Disease or Clinical Signs | Interrelationships or Toxicity |
|---|---|---|---|---|
Cobalt (Co) | 0.2 | Component of vitamin B12 (ruminants only) | Megaloblastic anemia; enzootic marasmus (ketosis), decreased growth, body weight | Cobalt needs to be supplied to rumen microbes; toxicosis unlikely |
Copper (Cu) | 9 | Hemoglobin synthesis; bone formation; pigmentation; myelin formation; oxidation-decrease reactions; immune function | Achromotrichia; neonatal ataxia (swayback); infertility; hypochromic-microcytic anemia; profuse diarrhea; immune failure | High iron, zinc, and selenium decrease availability; excess molybdenum and sulfur inhibit utilization and storage in ruminants; toxicity variable in species, results in hemolytic crisis |
Iodine (I) | 0.5 | Thyroxine formation | Goiter; stillbirths, infertility; malformations, hairless neonates; immune failure | Dietary goitrogens inhibit function; high intakes decrease uptake efficiency; toxic intake results in goiter, fetal malformations |
Iron (Fe) | 50–100 | Cellular respiration, oxygen transport | Hypochromic-microcytic anemia; easily fatigued | High calcium may decrease absorption; copper status influences metabolism; excessive amounts are pro-oxidants; interferes with copper metabolism |
Manganese (Mn) | 30–40 | Bone formation; activator of enzymes with carbohydrate, lipid, and protein metabolism | Crooked calf disease; impaired fertility; poor growth; impaired immune function | Excess calcium and phosphorus decrease availability; toxicosis unlikely; may interfere with other minerals |
Selenium (Se) | 0.3 | Antioxidant—cellular iodine function, immune function | White muscle disease; acute death; retained placenta; impaired immune function | Sulfate, acid soil conditions interferes with selenium availability; acute toxicosis from parenteral products— respiratory collapse |
Zinc (Zn) | 28–60 | Component of > 90 enzymes influencing all aspects of metabolism immune function | Anorexia; parakeratosis; poor skin and hair formation; male infertility; impaired immune function | High calcium, phytates or oxalates bind zinc; high zinc interferes with copper metabolism |
Trace mineral nutritional status is best assessed by determining hepatic mineral concentration, as the liver is the primary storage site for all trace minerals except iodine. Blood analysis can be used for some trace minerals; however, one must understand potential confounding issues limiting interpretation.
Selenium status of cattle can be accurately assessed from blood or serum concentrations. Whole blood concentrations of 120–250 ng/mL or serum concentrations of 70–100 ng/mL in adult cattle indicate adequate selenium status.
Copper status of cattle can be assessed best from liver and from serum copper concentrations only when multiple samples are collected. Liver concentrations < 50 mg/kg dry tissue or serum concentrations < 0.3 mcg/mL indicate inadequate copper status.
Normal serum zinc concentrations are 0.7–1.3 mcg/mL. Concentrations < 0.4 mcg/mL are considered deficient. Blood sampling for zinc determination should use special trace mineral tubes (with a royal blue stopper) to prevent contamination from blood contacting the zinc-containing lubricant used for rubber-based stoppers.
Adequate serum iron concentrations are 110–150 mcg/dL. However, these concentrations decrease rapidly with inflammatory disease, and such changes in serum iron concentrations should not be interpreted as being due to dietary deficiency.
Vitamins
Vitamin A Requirements of Dairy Cattle
The biological form of vitamin A, retinol, does not exist in any plant material, so there is no vitamin A in natural diets for dairy cattle. Vitamin A activity from natural sources comes primarily from beta carotene, which is found in plants and is particularly abundant in fresh forages. Beta carotene is labile: its concentrations in forages are not constant but diminish with time in storage. Therefore, measurement of beta carotene concentrations in feeds is not practical and is seldom done. Recommended vitamin A consumption rates for various classes of cattle are based on providing supplemental vitamin A, which is derived from commercial sources:
Adult cows (lactating and dry)—110 IU/kg body weight, which is approximately 4,400 IU/kg dry diet. If milk production exceeds 35 kg/day, 1,000 IU/kg milk produced beyond 35 kg are added.
Growing heifers—80 IU/kg body weight, which is approximately 2,500 IU/kg dry diet
Vitamin A requirements are increased with low-forage diets, high–corn silage diets, poor-quality forages, and infection.
Vitamin A deficiency is associated initially with night blindness, followed by poor growth, poor coats, and suppressed immunity. In adult cattle, vitamin A deficiency is associated with retained placentas and impaired fertility.
Calves are born with low body stores of vitamin A and depend on colostrum consumption to supply hepatic vitamin A. Minimal fetal vitamin A status may cause stillbirth and weak calves. The NASEM 2021 report recommends dietary vitamin A concentrations for young calves at approximately 9,000 IU/kg diet dry matter. Most milk replacer diets have substantially higher concentrations of vitamin A, possibly because vitamin A requirements may be increased by infectious diseases, especially those affecting the respiratory or enteric epithelium.
The vitamin A status of cattle may be assessed via serum or hepatic vitamin A concentrations. The liver stores vitamin A for release during periods of insufficient dietary intake, making liver the ideal tissue for nutritional assessment. For adult cattle receiving diets with recommended supplemental vitamin A concentrations, hepatic vitamin A concentrations are 300–1,100 mg/kg dry tissue (expressed as retinol). Clinical signs of vitamin A deficiency do not occur until these reserves have been substantially depleted. Adequate serum vitamin A concentrations in adult cattle are 225–500 ng/mL, usually decreasing to approximately 150 ng/mL within 1 week after calving.
Vitamin D Requirements of Dairy Cattle
Vitamin D plays a central role in homeostasis of calcium and phosphorus.New research has highlighted the role of vitamin D in immune cell function and gut immunity (8, 9, 10). Vitamin D3 (cholecalciferol) can be formed by the solar irradiation of skin or provided in the diet. A newer dietary vitamin D supplement provides the 25-hydroxycholecalciferol compound. Cattle can utilize vitamin D2 that is produced from solar irradiation of forages. However, natural vitamin D as a source is considered unreliable, and vitamin D requirements are based on recommendations for supplementing diets. The recommended rate of vitamin D supplementation for heifers and dry dairy cows is 30 IU/kg body weight. Adequate intake of vitamin D for lactating cows has been increased to 40 IU/kg body weight (approximately 1,100 IU/kg dry matter).
Vitamin D status can be assessed via serum concentrations of 25-hydroxycholecalciferol. Adequate concentrations are 20–50 ng/mL, with concentrations < 10 ng/mL indicating deficiency. Concentrations > 300 ng/mL indicate potential toxicosis.
Vitamin E Requirements of Dairy Cattle
Natural sources of vitamin E are derived from plant oils and are designated RRR-alpha-tocopherol or d-alpha-tocopherol, based on stereoisomer characteristics of their chemical structure. Vitamin E is present in relatively high concentrations in fresh forages. Thus, cattle receiving pasture or fresh-cut forages may require little vitamin E supplementation. In contrast, vitamin E degrades in stored forages, so dairy cattle on typical confinement-reared diets require supplemental vitamin E.
Vitamin E functions to protect cellular membranes from oxidative damage. Clinical manifestations of deficiency include nutritional myopathy (white muscle disease) in young calves and diseases in older cattle including retained placenta and increased susceptibility to environmental mastitis. Recommended rates of vitamin E intake vary based on physiological stage:
Far off dry period—1.6 IU/kg body weight (approximately 1,100 IU/day)
Close-up dry period (last 3 weeks)—3.0 IU/kg body weight, which is approximately 2,100 IU/kg dry matter
Heifers and lactating cows—0.8 IU/kg body weight, which is approximately 30 IU/kg dry matter
Higher vitamin E supplementation rates (4,000 IU/day close-up dry period; 2,000 IU/day early lactation) are occasionally provided when environmental mastitis is a particular problem. Vitamin E is essentially nontoxic, and there is little risk of oversupplementation.
Vitamin E supplements may be natural or synthetic. Synthetic supplements are designated all-rac-alpha-tocopherol, or dl-alpha-tocopherol. The natural source supplements seem to have much greater biological activity.
Serum concentrations may be used to assess vitamin E status in dairy cattle. Serum concentrations of 2–4 mcg/mL are generally adequate. Cow serum concentrations < 3 mcg/mL are associated with higher susceptibility to mastitis. However, in addition to vitamin E nutritional status, these concentrations are influenced by the total concentration of serum lipid, with higher serum lipid concentrations resulting in higher vitamin E concentrations. Serum lipids are generally low in late gestation and high in the period of peak feed intake. To compensate for this fluctuation, serum vitamin E concentrations are sometimes expressed as a ratio, with some serum lipid component, such as cholesterol or triglyceride, used as the denominator.
Other Vitamin Requirements of Dairy Cattle
Supplementation of other compounds such as vitamin K, B-complex vitamins, and vitamin C is not typical in most ruminant diets. Cows, unlike humans and primates, synthesize vitamin C adequately. Vitamin K and the B-complex vitamins are believed to be sufficiently synthesized by rumen and intestinal microbes to meet the cow’s needs.
Some vitamins can be provided via natural feed ingredients. There are cases in which B-vitamins might be supplemented, primarily during disease conditions and especially in conditions of altered rumen fermentation. Supplementation of rumen-protected B-vitamins (specifically, biotin, niacin, and choline) can benefit health and performance of high-producing dairy cattle; see the table ).
Summary of Dairy Cattle Feeding Recommendations From NASEM 2021
The NASEM committee undertook many challenges in revising and improving upon feeding recommendations for dairy cattle. A new approach to defining a “requirement” was incorporated in the report, which better represents the data support for the requirement (see the table ). Many of the trace mineral and vitamin requirements were defined as "adequate intake," indicating a lack of definitive information to adequately define a requirement. Energy requirement calculations were further refined with no significant change in values. However, the maintenance energy requirement was increased by 25%. This increase accounts for the majority of the increase in the dry cow energy requirement.
The use of metabolizable protein (MP) to define dairy cow amino acid requirements over crude protein is more strongly supported in this report. Metabolizable protein refers to all protein available for digestion in the small intestine, including microbial protein and protein not digested in the rumen. Rather than considering only the crude protein of a diet, MP is a more comprehensive description of the protein available to the cow. Improved characterization of feed protein fractions and amino acid composition is provided in compositional tables in the report. The modeling of protein supply has been modified from the NRC 2001 report, resulting in a complete modification of essential amino acid (EAA) recommendations and a focus on balancing diets for EAAs rather than MP. Although many improvements have been made to the MP prediction model, the report highlights much-needed research areas, especially relative to amino acid supply and utilization efficiency.
Similar to the modeling approach with MP focusing on EAAs, the feeding of fat focuses on compositional fatty acids. The new model of fat utilization allows for direct entry of fatty acid measurements rather than using ether extract. A new measure of dietary fiber has also been introduced—namely, physically adjusted neutral detergent fiber (paNDF). This parameter approach integrates the associative effects of dietary starch and forage type on the ability of NDF particle size to maintain a healthy rumen pH.
Refinements to macromineral requirements were made based on source bioavailability and significant mineral interactions. Among the trace minerals and fat-soluble vitamins, there were some notable changes to cobalt, copper, manganese, and zinc compared to the NRC 2001 report. Vitamin A and D requirements were increased for lactating cows, whereas vitamin E was increased in the close-up dry cow compared to NRC 2001.
NASEM Requirement Definitions
Estimated average requirement (EAR) : This is the nutrient amount needed to meet the requirement in half the healthy individuals in a particular life stage and gender group. This is the most robust requirement value, because it is based on many studies that determined variation in nutrient intake. |
Recommend dietary allowance (RDA): This requirement accounts for a greater proportion of the population in meeting the nutrient needs for nearly all healthy individuals in a particular life stage and gender group. It is defined as the EAR + 2 standard deviations; if EAR variation is unknown, then RDA = EAR × 1.2. |
Adequate intake (AI): Average daily nutrient intake that should meet or exceed requirements of specific groups. This value is determined by a panel of experts interpreting limited experimental data. Nutrients defined by this value have limited experimental data with high variation, and the EAR or RDA cannot be determined. |
Tolerable upper intake level (UL) : Highest average daily nutrient intake that is not likely to pose an adverse health risk to nearly all individuals in a population. This value is similar to the maximum tolerable level (MTL). Maximum tolerable level is defined as the highest dietary amount of a mineral that can be fed for a brief (up to 3 months) period without causing toxicosis. |
Substantial changes were made in addressing nutrient requirements of the nonlactating pregnant cow.
Energy and protein requirements were based on a continuous growth model of the conceptus (uterus, placenta, and fetus; see ). This approach more accurately describes the underlying gestational biology; however, it makes formulating a diet to match the changing requirements difficult. Unfortunately, insufficient data remain to include energy and protein required for mammary growth and colostrogenesis.
The model to predict DMI for the dry cow now accounts for dietary NDF content and its impact on decreasing intake capacity (see ).


Gestational conceptus growth and associated metabolizable energy (ME) and metabolizable protein (MP) requirements to support gestation (assuming cow body weight of 725 kg and calf birth weight of 45 kg). Based on data from Nutrient Requirements of Dairy Cattle, 2021.
Courtesy of Dr. Robert Van Saun.


Predicted dry matter intake (DMI; percent of live body weight) for nonlactating pregnant cows based on week prior to calving and dietary neutral detergent fiber (NDF) content compared to the NRC 2001 model. Adapted from Nutrient Requirements of Dairy Cattle, 2021.
Courtesy of Dr. Robert Van Saun.
Another improvement can be seen in revised models for the growing calf and heifer. Growth modeling was based on energy-allowable growth, which then directed protein requirements above maintenance. In contrast to the previous NRC 2001 report, prediction of retained energy is central to the new calf growth model in NASEM 2021. Studies documenting protein and fat composition of tissue deposited during calf growth provided additional data used to refine growth requirements. Calf mineral and vitamin requirements were provided on a factorial approach rather than a dietary composition.
Key Points
Substantial changes have been made in dairy cattle nutrition recommendations, as published in the 2021 report by the National Academies of Science, Engineering, and Medicine committee (formerly the National Research Council).
Balancing fiber and NFC/NSC fractions to optimize energy intake and rumen health is a challenging aspect of dairy nutrition.
Dietary fiber needs (paNDF) are influenced by forage neutral detergent fiber (NDF), dietary starch, forage fragility, and particle size distribution.
For More Information
National Academies of Science, Engineering, and Medicine (NASEM). Nutrient Requirements of Dairy Cattle. 8th rev ed. National Academies Press; 2021.
Van Saun RJ, ed. In: Dairy nutrition. Vet Clin North Am Food Anim Pract. 2014;30(3):487-804.
McNamara JP, Lucy MC, eds. In: Volume 100 special issue. J Dairy Sci. 2017;100(12):9892-10446.
Swecker WS Jr, Van Saun RJ, eds. In: Ruminant trace mineral nutrition. Vet Clin North Am Food Anim Pract. 2023;39(3):385-558.
Nolan J and Primary Industries Standing Committee. Nutrient Requirements of Domesticated Ruminants. CSIRO Publishing; 2007.
Pond WG, Church DB, Pond KR, Schoknecht PA. Basic Animal Nutrition and Feeding. John Wiley & Sons; 2004.
References
Goff JP, Koszewski NJ. Comparison of 0.46% calcium diets with and without added anions with a 0.7% calcium anionic diet as a means to reduce periparturient hypocalcemia. J Dairy Sci. 2018;101(6):5033-5045. doi:10.3168/jds.2017-13832
Glosson KM, Zhang X, Bascom SS, Rowson AD, Wang Z, Drackley JK. Negative dietary cation-anion difference and amount of calcium in prepartum diets: effects on milk production, blood calcium, and health. J Dairy Sci. 2020;103(8):7039-7054. doi:10.3168/jds.2019-18068
Zhang X, Glosson KM, Bascom SS, Rowson AD, Wang Z, Drackley JK. Metabolic and blood acid-base responses to prepartum dietary cation-anion difference and calcium content in transition dairy cows. J Dairy Sci. 2022;105(2):1199-1210. doi:10.3168/jds.2021-21191
Graef GM, Kerwin AL, Ferro LN, et al. Effect of prepartum dietary cation-anion difference strategy and level of dietary calcium on postpartum blood calcium status and milk production of multiparous Holstein cows. J Dairy Sci. 2025;108(8):8332-8349. doi:10.3168/jds.2024-25783
Kerwin AL, Ryan CM, Leno BM, et al. Effects of feeding synthetic zeolite A during the prepartum period on serum mineral concentration, oxidant status, and performance of multiparous Holstein cows. J Dairy Sci. 2019;102(6):5191-5207. doi:10.3168/jds.2019-16272
Thilsing-Hansen T, Jorgensen, RJ, Enemark JMD, Larsen T. The effect of zeolite A supplementation in the dry period on periparturient calcium, phosphorus, and magnesium homeostasis. J Dairy Sci. 2002;85(7):1855-1862. doi:10.3168/jds.S0022-0302(02)74259-8
Frizzarini WS, Campolina JP, Vang AL, et al. Mechanisms by which feeding synthetic zeolite A and dietary cation-anion difference diets affect feed intake, energy metabolism, and milk performance: part II. J Dairy Sci. 2024;107(7):5204-5221. doi:10.3168/jds.2024-24057
Eder K, Grundmann SM. Vitamin D in dairy cows: metabolism, status and functions in the immune system. Arch Anim Nutr. 2022;76(1):1-33. doi:10.1080/1745039X.2021.2017747
Vieira-Neto A, Poindexter MB, Nehme Marinho M, et al. Effect of source and amount of vitamin D on function and mRNA expression in immune cells in dairy cows. J Dairy Sci. 2021;104(10):10796-10811. doi:10.3168/jds.2021-20284
Golder HM, McGrath J, Lean I. Effect of 25-hydroxyvitamin D3 during prepartum transition and lactation on production, reproduction, and health of lactating dairy cows. J Dairy Sci. 2021;104(5):5345-5374. doi:10.3168/jds.2020-18901





