



Calcium and Phosphorus Imbalances
A deficiency of either calcium or phosphorus in the diet of young growing birds results in abnormal bone development, even when the diet contains adequate vitamin D3. A deficiency of either calcium or phosphorus results in lack of normal skeletal calcification. Rickets is seen mainly in growing birds, whereas calcium deficiency in laying hens results in reduced shell quality and subsequently osteoporosis. This depletion of bone structure causes a disorder commonly referred to as cage layer fatigue since it was first seen with the introduction of cage housing systems some 60 years ago. The condition is rarely seen in noncage systems, although this situation is confounded with lower egg production. Over time, excessive mobilization of bone calcium to overcome a dietary deficiency causes structural bone to erode, and it is then unable to support the weight of the hen.
Rickets
Rickets occurs most commonly in young meat birds; the main characteristic is inadequate bone mineralization. Calcium deficiency at the cellular level is the main cause, although feeding a diet deficient or imbalanced in calcium, phosphorus, or vitamin D3 can also induce this problem. Young broilers and turkey poults can exhibit lameness at ~10–14 days of age. Their bones are rubbery, and the ribs become flattened and beaded at the attachment of the vertebrae. Rachitic birds exhibit a disorganized cartilage matrix, with an irregular vascular penetration. There is an indication of impaired metabolism of collagen precursors such as hyaluronic acid and desmosine.
Rickets is not caused by a failure in the initiation of bone mineralization but rather by impairment of the early maturation of this process. There is often an enlargement of the ends of the long bones, with a widening of the epiphyseal plate. A determination of whether rickets is due to deficiencies of calcium, phosphorus, or vitamin D3 or to an excess of calcium (which induces a phosphorus deficiency) may require analysis of blood phosphorus levels and investigation of parathyroid activity.
In most field cases of rickets, a deficiency of vitamin D3 is suspected. This can be due to simple dietary deficiency, inadequate potency of the D3 supplement, or other factors that reduce the absorption of vitamin D3.
Rickets can best be prevented by providing adequate levels and potency of vitamin D3 supplements and by ensuring the diet is formulated to optimally use all fat-soluble compounds. Young birds have limited ability to digest saturated fats, and these undigested compounds can complex with calcium to form insoluble soaps, leading to an induced deficiency of calcium. Again, this situation cannot be diagnosed through diet assay for calcium but rather through excreta assay of this mineral.
Diets must also provide a correct balance of calcium to available phosphorus. For this reason, ingredients notoriously variable in their content of these minerals, such as animal proteins, should be used with extra caution.
In recent years, the use of 25(OH)D3 has become very common as a partial replacement for vitamin D3, with reports of greatly reduced incidence of rickets, especially in turkey poults. This metabolite has the same structure as that naturally produced in the liver of birds in the first step of conversion of vitamin D3 to 1,25(OH)2D3, which is the active form of the vitamin. The commercial form of 25(OH) D3 is therefore especially useful if normal liver metabolism is compromised in any way, such as occurs with mycotoxins or other natural toxins in the feed that potentially impair liver metabolism.
Tibial Dyschondroplasia (Osteochondrosis)

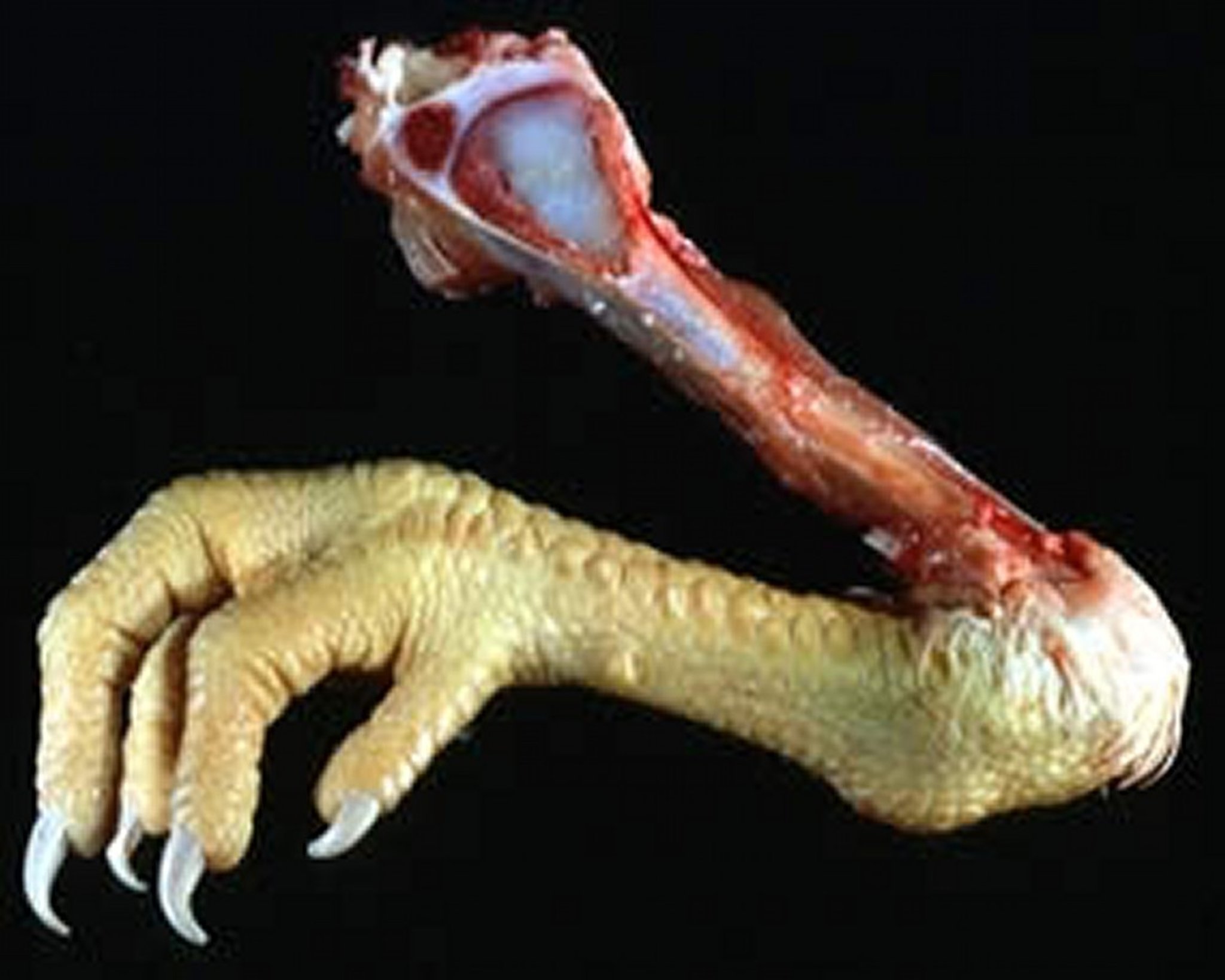
Cartilaginous core in the area of the growth plate in a young chicken with tibial dyschondroplasia.
Courtesy of Dr. Jean Sander.

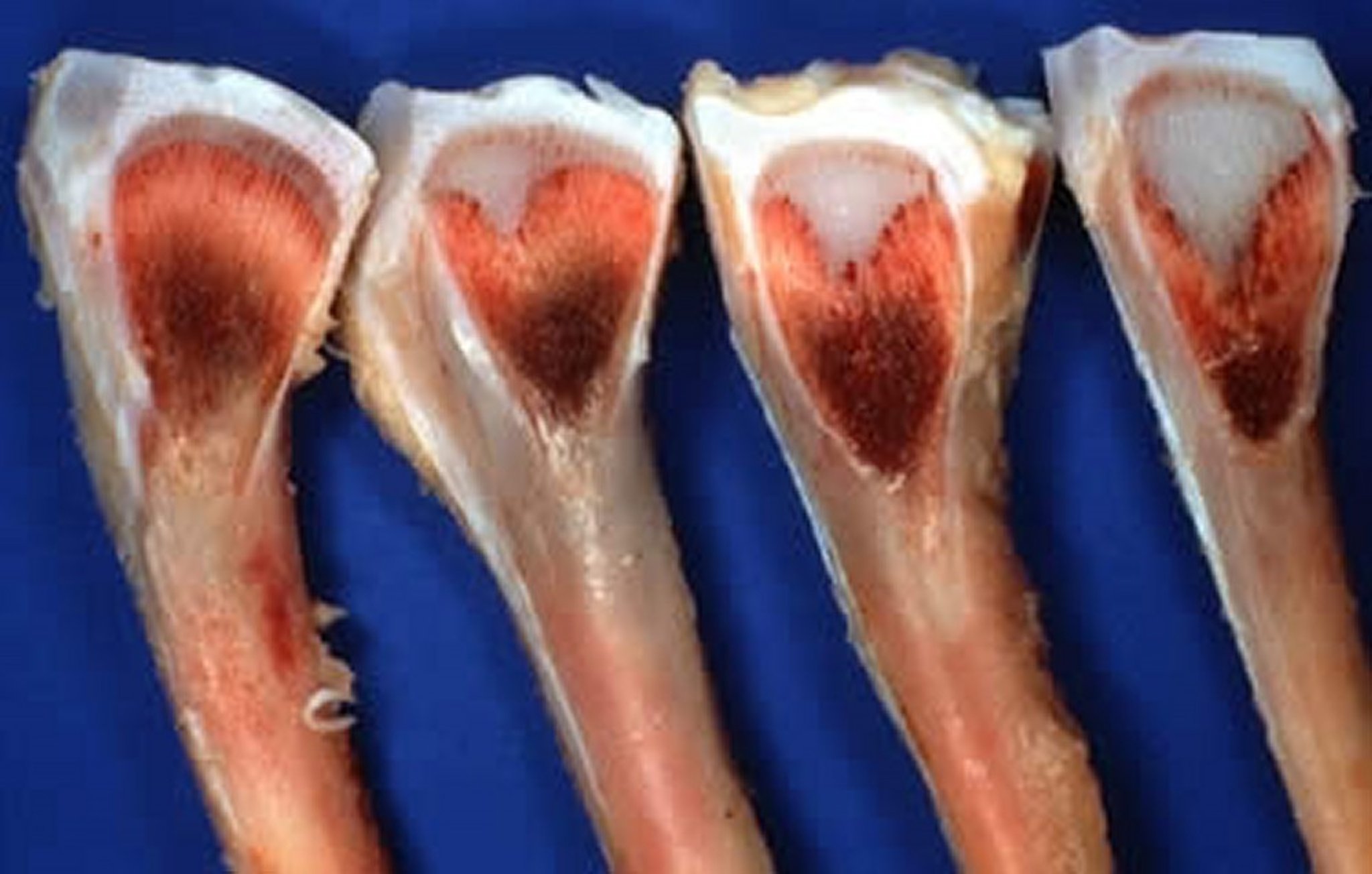
Varying degrees of severity of tibial dyschondroplasia, with a normal growth plate on the left for comparison.
Courtesy of Dr. Jean Sander.
Tibial dyschondroplasia is characterized by an abnormal cartilage mass in the proximal head of the tibiotarsus. It has been seen in all fast-growing types of meat birds but is most common in broiler chickens. Regardless of diet or environmental conditions, fast versus slow growth rate seems to at least double the incidence of tibial dyschondroplasia.
Signs can occur early but more usually are not initially seen until 14–25 days of age. Birds are reluctant to move, and when forced to walk, do so with a swaying motion or stiff gait. Tibial dyschondroplasia results from disruption of the normal metaphyseal blood supply in the proximal tibiotarsal growth plate, where the disruption in nutrient supply means the normal process of ossification does not occur. The abnormal cartilage is composed of severely degenerated cells, with cytoplasm and nuclei appearing shrunken. Affected cartilage contains less protein and less DNA.
The exact cause of tibial dyschondroplasia is unknown. Incidence can be altered through genetic selection and is likely affected by a major sex-linked recessive gene. Imbalance of dietary electrolytes, and particularly high levels of chloride relative to other dietary cations, seems to be a major contributor in many field outbreaks. More tibial dyschondroplasia is also seen when the level of dietary calcium is low relative to that of available phosphorus, or more commonly when diet phosphorus is high relative to calcium.
Treatment involves dietary adjustment of the calcium:phosphorus ratio and by achieving a dietary electrolyte balance of ~240 mEq/kg. Dietary changes rarely result in complete recovery. Tibial dyschondroplasia can be prevented by tempering growth rate; however, programs of light or feed restriction must be considered in relation to economic consequences of reduced growth rate. There is evidence that replacement of some of the dietary vitamin D3 with metabolites such as 1,25(OH)2D3 improves chondrocyte differentiation and hence limits occurrence of this skeletal disorder.
Cage Layer Fatigue
High-producing laying hens maintained in cages sometimes show paralysis during and just after the period of peak egg production due to a fracture of the vertebrae that subsequently affects the spinal cord. The fracture is caused by an impaired calcium flux related to the high output of calcium in the eggshell.
Layers are capable of early egg production exceeding 95% for at least 6 months, which places even more pressure on maintenance of adequate calcium flux between the diet, the skeleton, and the oviduct. As the hen ages, medullary bone is replaced on a daily basis as the demands for calcium diminish greatly immediately after oviposition and before the start of shell calcification of the next egg. However, structural (cortical and trabecular) bone is not replaced while the hen is in production. Over time, the medullary bone, which initially forms a protective layer over the structural bone, becomes more diffuse in the medullary cavity. This exposes more structural bone surface area to the activity of osteoclasts, and over time, osteoporosis can develop. The condition is rarely seen in floor-housed birds, suggesting that reduced activity within the cage is a predisposing or associated factor.
Affected birds are invariably found on their sides in the back of the cage. At the time of initial paralysis, birds appear healthy and often have a shelled egg in the oviduct and an active ovary. Death occurs from starvation or dehydration, because the birds simply cannot reach feed or water. Affected birds can recover if moved to the floor. A high incidence of cage layer fatigue can be prevented by ensuring the normal weight-for-age of pullets at sexual maturity and by giving pullets a high-calcium diet (minimum 4% calcium) for at least 7 days before first oviposition.
Older caged layers are also susceptible to bone breakage during removal from the cage and transport to processing. Bone strength cannot practically be improved during the production cycle without adverse consequences to other economically important traits such as eggshell quality. Cage layer fatigue is undoubtedly related to high, sustained egg output and associated clutch lengths of 200–230 eggs laid on successive days.
Diets must provide adequate quantities of calcium and phosphorus to prevent deficiencies. However, feeding diets that contain > 2.5% calcium during the immature growing period (< 16 weeks) produces a high incidence of nephritis, visceral gout, calcium urate deposits in the ureters, and sometimes high mortality, especially in the presence of infectious bronchitis virus.
Eggshell strength can be maintained, and the inevitable loss of bone strength can be mitigated, by feeding 65%–70% of the dietary calcium supplement in the form of coarse limestone (2–4 mm in diameter), with the remainder as fine particle limestone. Offering the coarse supplement permits the birds to satisfy their requirements when they need it most, allowing the coarse material to be retained in the gizzard where the calcium can be absorbed continually and especially at night when the bird is not feeding and when peak demand for shell calcification occurs.
A readily available calcium and/or calcium phosphate supplement is often effective if started very soon after paralysis is first observed. Although these supplements may be advantageous to afflicted layers, they are not ideal for the regular birds (95%–98%) in the flock; therefore, decisions regarding treatment are often influenced by the severity of the condition and the proportion of the flock affected.
Manganese Deficiency
A dietary manganese deficiency in immature chickens and turkeys is one of the potential causes of perosis and chondrodystrophy and also the production of thin-shelled eggs and poor hatchability in mature birds (see also ).
The most dramatic classic effect of manganese deficiency syndrome is perosis, characterized by enlargement and malformation of the tibiometatarsal joint, twisting and bending of the distal end of the tibia and the proximal end of the tarsometatarsus, thickening and shortening of the leg bones, and if severe, slippage of the gastrocnemius tendon from its condyles. Increased intakes of calcium and/or phosphorus will aggravate the condition because of reduced absorption of manganese via the action of precipitated calcium phosphate in the intestinal tract. In laying hens, reduced egg production, markedly reduced hatchability, and eggshell thinning are often noted.
A manganese-deficient breeder diet can result in chondrodystrophy in chick embryos. This condition is characterized by shortened, thickened legs and shortened wings. Other signs can include a parrot beak brought about by a disproportionate shortening of the lower mandible, globular contour of the head due to anterior bulging of the skull, edema occurring just above the atlas joint of the neck and extending posteriorly, and protruding of the abdomen due to unassimilated yolk. Growth is also reduced, and development of down and feathers is retarded. A manganese-deficient chick has a characteristic star-gazing posture, because the physiology of the inner ear becomes defective.
Deformities cannot be corrected by feeding more manganese. Effects of manganese deficiency on egg production are fully corrected by feeding a diet that contains at least 30–40 mg of manganese/kg, provided the diet does not contain excess calcium and/or phosphorus. There is an indication of the need for Fe2+ ions as well as manganese to correct the deficiency, although most commercial poultry diets contain a surfeit of iron.
Iron and Copper Deficiencies
A deficiency of either iron or copper in poultry can lead to anemia. Iron deficiency causes a severe anemia, with a reduction in PCV. In color-feathered strains, there is also loss of pigmentation in the feathers. The birds’ requirements for synthesis of red blood cells takes precedence over metabolism of feather pigments, although if a fortified diet is introduced, all subsequent feather growth is normal and lines of demarcation on the feathers are part of diagnosis. Iron may be needed not only for the red feather pigments, which contain iron, but also to function in an enzyme system involved in the pigmentation process. Ochratoxin at 4–8 mcg/g diet also causes an iron deficiency characterized by hypochromic, microcytic anemia. Aflatoxin also reduces iron absorption.
Young chicks become lame within 2–4 weeks when fed a copper-deficient diet. Bones are fragile and easily broken, the epiphyseal cartilage becomes thickened, and vascular penetration of the thickened cartilage is markedly reduced. These bone lesions resemble the changes noted in birds with a vitamin A deficiency. Copper is required for cartilage formation, and certain antinutrients such as some grain fumigants have been shown to impact skeletal development, likely via interaction with copper metabolism. Copper-deficient chickens may also display ataxia and spastic paralysis.
Copper deficiency in birds, and especially in turkeys, can lead to rupture of the aorta. The biochemical lesion in the copper-deficient aorta is likely related to failure to synthesize desmosine, the cross-link precursor of elastin. The lysine content of elastin in copper-deficient birds is three times that seen in control birds, suggesting failure to incorporate lysine into the desmosine molecule. In field cases of naturally occurring aortic rupture, many birds have < 10 ppm copper in the liver, compared with 15–30 ppm normally seen in birds of comparable age. High levels of sulfate ions, molybdenum, and ascorbic acid can reduce liver copper levels.
Iodine Deficiency
Iodine deficiency results in a decreased output of thyroxine from the thyroid gland, which in turn stimulates the anterior pituitary to produce and release increased amounts of thyroid stimulating hormone (TSH). This increased production of TSH results in subsequent enlargement of the thyroid gland, usually termed goiter. The enlarged gland results from hypertrophy and hyperplasia of the thyroid follicles, which increases the secretory surface of the follicles.
Lack of thyroid activity or inhibition of the thyroid by administration of thiouracil or thiourea causes hens to cease laying and become obese. It also results in the growth of abnormally long, lacy feathers. Administration of thyroxine or iodinated casein reverses the effects on egg production, with eggshell quality returning to normal. The iodine content of an egg is markedly influenced by the hen’s intake of iodine. Eggs from a breeder hen fed an iodine-deficient diet will exhibit reduced hatchability and delayed yolk sac absorption. Unimproved rapeseed meal can contain sufficient levels of goitrogens that cause thyroid enlargement in young birds. Iodine deficiency in poultry can be avoided by supplementing the feed with as little as 0.5 mg of iodine/kg, although a level of 2–3 mg/kg is more commonly provided to sustain good feathering in fast-growing birds.
Magnesium Deficiency
Natural feed ingredients are rich in magnesium; thus, deficiency is rare and magnesium is never specifically used as a supplement to poultry diets. Newly hatched chicks fed a diet totally devoid of magnesium live only a few days. They grow slowly, are lethargic, and often pant and gasp. When disturbed, they exhibit brief convulsions and become comatose, which is sometimes temporary but often fatal. Mortality is quite high on diets only marginally deficient in magnesium, even though growth of survivors may approach that of control birds.
A magnesium deficiency in laying hens results in a rapid decline in egg production, hypomagnesemia, and a marked withdrawal of magnesium from bones. Egg size, shell weight, and the magnesium content of yolk and shell are decreased. Increasing the dietary calcium of laying hens accentuates these effects. Magnesium seems to play a central role in eggshell formation, although it is not clear whether there is a structural need or whether magnesium simply gets deposited as a cofactor along with calcium.
Magnesium requirements for most classes of chickens seem to be ~500–600 ppm, a level usually achieved with contributions from natural feed ingredients.
Potassium, Sodium, and Chloride Deficiencies
Although requirements for potassium, sodium, and chloride have been clearly defined, it is also important to maintain a balance of these and all other electrolytes in the body. Often termed electrolyte balance or acid-base balance, the effects of deficiency of any one element are often a consequence of alteration to this important balance as it affects osmoregulation.
Simple Deficiency
A deficiency of chloride causes ataxia with classic signs of nervousness, often induced by sudden noise or fright. The main sign of hypokalemia is an overall muscle weakness characterized by weak extremities, poor intestinal tone with intestinal distention, cardiac weakness, and weakness and ultimately failure of the respiratory muscles. Hypokalemia is apt to occur during severe stress. Plasma protein is increased, causing the kidney, under the influence of adrenocortical hormone, to discharge potassium into the urine. During adaptation to the stress, blood flow to the muscle gradually improves, and the muscle begins uptake of potassium. As liver glycogen is restored, potassium returns to the liver.
Birds fed a diet low in both protein and potassium or that are starving grow slowly but do not show a potassium deficiency. Potassium derived from catabolized tissue protein replaces that lost in the urine. The ratio of potassium to nitrogen in urine is relatively constant and is the same as that found in muscle. Thus, tissue nitrogen and potassium are released together from the catabolized tissue.
A deficiency of sodium leads to a lowering of osmotic pressure and a change in acid-base balance in the body. Cardiac output and blood pressure both decrease, PCV increases, elasticity of subcutaneous tissues decreases, and adrenal function is impaired. This leads to an increase in blood uric acid levels, which can result in shock and death. A less severe sodium deficiency in chicks can result in retarded growth, soft bones, corneal keratinization, impaired food utilization, and a decrease in plasma volume. In layers, reduced egg production, poor growth, and cannibalism may be noted. A number of diseases can result in sodium depletion from the body, such as GI losses from diarrhea or urinary losses due to renal or adrenal damage.
Electrolyte Imbalance
Electrolyte balance is commonly described by the simple formula of Na + K – Cl expressed as mEq/kg of diet. An overall dietary balance of 240–260 mEq/kg is generally considered optimal for normal physiologic function. The buffering systems in the body ensure the maintenance of near normal physiologic pH, preventing electrolyte imbalance. The primary role of electrolytes is in maintenance of body water and ionic balance. Thus, requirements for elements such as sodium, potassium, and chloride cannot be considered individually, because it is the overall balance that is important. Electrolyte balance, also referred to as acid-base balance, is affected by three factors:
the balance and proportion of these electrolytes in the diet
endogenous acid production
the rate of renal clearance
In most situations, the body maintains a normal balance between cations and anions in the body such that physiologic pH is maintained. If there is a shift toward acid or base conditions, metabolic processes return the body to a normal pH. Actual electrolyte imbalances are rare, because regulatory mechanisms must sustain optimal cellular pH and osmolarity. Electrolyte balance can therefore more correctly be described as the changes that necessarily occur in the body processes to achieve normal pH. In extreme situations, such modifications in regulatory mechanisms seem to adversely affect other physiologic systems, and they produce or accentuate potentially debilitating conditions.
Electrolyte imbalance causes a number of metabolic disorders in birds, most notably tibial dyschondroplasia and respiratory alkalosis in layers. The unusual development of a cartilage plug at the growth plate of the tibia can be induced by a number of factors, although its incidence can be greatly increased by metabolic acidosis induced by feeding products such as NH4Cl. Tibial dyschondroplasia seems to occur more frequently when the diet contains an excess of sodium relative to potassium, along with very high chloride levels. The latter situation is most easily remedied by substitution of sodium bicarbonate for sodium chloride in the diet.
Overall electrolyte balance is always important but is most critical when chloride or sulfur levels are high. With low dietary chloride levels, there is often little response to the manipulation of electrolyte balance; however, when dietary chloride levels are high, it is critical to make adjustments to the dietary cations to maintain overall balance. Alternatively, chloride levels can be reduced, although chickens have requirements of ~0.12%–0.15% of the diet, and deficiency signs will develop with dietary levels < 0.12 %.
Sodium content of drinking water can have a meaningful impact on total sodium intake of the bird. When drinking water contains > 300 ppm of sodium, it may be necessary to reduce sodium levels in the diet. Supplementation of exogenous phytase impacts electrolyte balance. This commonly used enzyme is intended to reduce dependence on supplemental inorganic phosphorus, but it concomitantly reduces renal excretion of sodium. Diets, therefore, need less supplemental sodium when they contain phytase.
Selenium Deficiency
A deficiency of selenium in growing chickens causes exudative diathesis. Early signs of unthriftiness and ruffled feathers usually occur at 3–6 weeks of age, depending on the degree of deficiency. The edema results in weeping of the skin, which is often seen on the inner surface of the thighs and wings. The birds bruise easily, and large scabs often form on old bruises. In laying hens, such tissue damage is unusual, but egg production, hatchability, and feed conversion are adversely affected.
The metabolism of selenium is closely linked to that of vitamin E, and signs of deficiency can sometimes be treated with either the mineral or the vitamin. Vitamin E can spare selenium in its role as an antioxidant, and so some selenium-responsive conditions can also be treated by supplemental vitamin E. In most countries, there are limits to the quantity of selenium that can be added to a diet; the upper limit is usually 0.3 ppm because of the potential for toxicity at relatively low levels.
The commonly used forms of selenium are sodium selenite and, more recently, organic selenium chelates. Feeds grown on high-selenium soils are sometimes necessarily used in poultry rations and are good sources of selenium. Fish meal and dried brewer’s yeast are also rich in available selenium.
Zinc Deficiency
Zinc requirements and signs of deficiency are influenced by dietary ingredients. In semipurified diets, it is difficult to show a response to zinc levels much above 25–30 mg/kg diet, whereas in practical corn-soybean meal diets, requirement values are increased to 60–80 mg/kg. Such variable zinc needs likely relate to phytic acid content of the diet, because this ligand is a potent zinc chelator. If phytase is used in diets, the need for supplemental zinc is reduced by up to 10 mg/kg diet.
In young chicks, signs of zinc deficiency include:
retarded growth
shortening and thickening of leg bones and enlargement of the hock joint
scaling of the skin (especially on the feet)
very poor feathering
loss of appetite
in severe cases, mortality
Although zinc deficiency can reduce egg production in aging hens, the most striking effects are seen in developing embryos. Chicks hatched from zinc-deficient hens are weak and cannot stand, eat, or drink. They have accelerated respiratory rates and labored breathing. If the chicks are disturbed, the signs are aggravated and the chicks often die. Retarded feathering and frizzled feathers are also found. However, the major defect is grossly impaired skeletal development. Zinc-deficient embryos show micromelia, curvature of the spine, and shortened, fused thoracic and lumbar vertebrae. Toes often are missing and, in extreme cases, the embryos have no lower skeleton or limbs. Some embryos are rumpless, and occasionally the eyes are absent or not developed.





